Introduction
Gull-billed
Terns (Sterna nilotica), Common Terns (S. hirundo), Black
Skimmers (Rynchops niger), and American Oystercatchers (Haematopus
palliatus) nest on storm-deposited shellpiles on salt marsh islands and on
barrier island beaches in the barrier island-lagoon complex that characterizes
the Eastern Shore of Virginia. Most shellpiles have low elevations, and may be
subject to flooding due to spring high tides and storm surges during the
breeding season (Erwin et al. 1998, Eyler et al. 1999). Total inundation of
shellpile habitats and marshes occurs more often than on barrier islands (B.R.
Truitt, TNC, pers. comm.). Populations of Gull-billed Terns, Common Terns, and
Black Skimmers nesting on Virginia’s barrier islands have declined by 95%, 84%,
and 86%, respectively, from 1975 to 1999 (Williams et al. 1990, B. Williams
unpubl. data). American Oystercatcher populations have declined by more than
50% on Virginia barrier islands over the last 20 years (Davis et al. 2001).
However, these numbers may not reflect population changes in all of Virginia,
since marsh island populations are not included in these surveys.
One of the most significant
threats to waterbirds in the Atlantic region is the decline in both the
quantity and quality of habitat (Parnell et al. 1988, Erwin et al. in press).
Because most of Virginia’s barrier island habitats are protected, they have not
experienced the same magnitude of human disturbance as other parts of the
Atlantic Coast (Erwin 1980). However, over the last 25 years, the distribution
of raccoons (Procyon lotor) and red foxes (Vulpes vulpes) has
increased on Virginia barrier islands while the number of beach-nesting tern
and skimmer colonies has decreased (Erwin et al. 2001). Shellpiles on salt
marsh islands provide a haven from mammalian predators, but flooding during the
breeding season may cause increased breeding failure (Burger and Lesser 1979,
Eyler et al. 1999). In addition, loss of tidal marshes due to human encroachment
has reduced available breeding and feeding habitat (Parnell et al. 1988).
Because of growing mammalian predator populations on barrier islands, and
frequent flooding on salt marsh shellpiles, safe nesting sites for waterbirds
are limited in coastal Virginia (Erwin et al. 2001). The reduction in the
quantity of suitable breeding habitat may continue if the predicted rise in sea
level, due to global atmospheric warming, increases flooding of coastal areas
(NAS 1987). A slight increase in sea level and/or higher frequency of storms
could increase the frequency of flooding on shellpiles and barrier island
beaches, and thus cause greater rates of breeding failure. Until recently the
primary risk to marine birds was human habitat destruction (Brown 1991, Nettleship
et al. 1994), but suitable breeding habitat for waterbirds may be even more
restricted in the next decades as a rising sea encroaches on nesting sites, and
mammalian predators continue to inhabit barrier islands beaches.
Because of habitat loss, expanding
mammalian populations, and rising sea levels, management of waterbird breeding
sites may become increasingly necessary. Habitat enhancement by manipulation of
nesting sites needs to be attempted as a method to reduce the frequency of
flooding, and to determine methods to develop and/or protect nesting sites that
will be suitable during times of higher sea levels. The overall objectives of
this project are to determine: (1) how four species of waterbirds differ in
their nest-site choices, (2) how biological and physical factors affect that
choice, (3) whether manipulation of habitat elevation influences nest-site
choice, and (4) whether manipulation of habitat elevation can improve
reproductive success.
Study Site
A
barrier island-lagoon complex extends about 100 km from Chincoteague Bay to
Kiptopeke Point along the eastern shore of the Delmarva Peninsula, Virginia
(Fig 1). Some salt marsh islands within the lagoons have storm-deposited oyster
shellpiles along marsh edges. The shellpiles have higher elevations than the
surrounding marsh, and vegetation is present on lower-lying areas of some
shellpiles. Wrack (dead vegetation mats of either Spartina spp. or Zostera
marina) deposited by high tides and storms often rings the shellpiles,
sometimes covering the ridges. Five shellpiles were chosen for this study based
on their use by nesting waterbirds in 2001 (R. Rounds, pers. obs.) and previous
years (Erwin et al. 1998). Although the highest portions of a shellpile may
remain above water during high tides or storm surges, many other parts of the
shellpile are inundated. The approximate edges of the high-water limit can be
determined from the lines of recent wrack deposition.
Species Descriptions
Gull-billed Terns, Common
Terns, and Black Skimmers frequently form mixed-species colonies along the
Atlantic Coast on marsh, shellpile and beach habitats (Burger and Lesser 1978,
Buckley and Buckley 1984, Humphrey 1990, Burger and Gochfeld 1991, Gochfeld and
Burger 1994, Parnell et al. 1995). Black Skimmers often select colony
sites based on the presence of other species, especially Common and Gull-billed
Terns (Erwin 1977a, Pius and Leberg 1997). The American Oystercatcher is the
only solitary nester that breeds on the shellpiles, though they will commonly
nest near or in tern colonies (Burger 1985).
Figure 1. Map of the eastern shore of
Virginia with locations of study shellpiles

Surveys of beach-nesting
waterbirds on Virginia barrier islands from 1975-1994 found that Gull-billed
Tern populations have declined by 95% (Williams et al. 1990, B. Williams
unpubl. data). Gull-billed Terns are considered a threatened species by the
Commonwealth of Virginia and the Virginia Natural Heritage Program ranks the
species as G5/S2 meaning it is common throughout its range, but rare in
Virginia (Byrd and Johnston 1991). Williams et al. (1990, B. Williams unpubl.
data) found that Common Terns had decreased in coastal Virginia from 1975-1994
by 84% although some of the decrease may be related to a shift of more than
3000 breeding pairs to the Hampton Roads Bridge Tunnel (Erwin et al. 2001).
Black Skimmer populations declined on Virginia barrier islands by 86% from
1975-1994 (Williams et al. 1990, B. Williams unpubl. data). The largest number
of breeding pairs of American Oystercatchers on the Atlantic Coast is found in
Virginia and this population increased from 1986 to 1993 by 300 pairs (Nol and
Humphrey 1994). However, more recent data indicate that populations have
declined by more than 50% from 1979 to 1999 on Virginia barrier islands (Davis
et al. 2001). Threats to all species include loss of habitat to human
encroachment, environmental contamination, human disturbance, predation, and
localized flooding (Erwin 1980, Buckley and Buckley 1984, Williams et al. 1990,
Byrd and Johnston 1991, Via et al.1992, Gochfeld and Burger 1994, Nol and
Humphrey 1994, Erwin et al. 2001). Because of the declining population of these
waterbird species, it is important to determine management methods to enhance
breeding success at colony sites in Virginia and elsewhere.
Habitat and Nest-site Selection
Habitat selection can be
defined as a series of decisions made by an individual resulting in the use of
one habitat in preference to other available habitats (Jones 2001). Because
habitat selection involves choices, it can be thought of as a behavioral
process in which habitats are assessed and then selected for their influence on
an individual’s survival and fitness (Fretwell 1972, Jones 2001). Fretwell
(1972) describes an “ideal free distribution” model for habitat selection in
which individuals are free to select the habitat most suitable to them and to
settle there. However, as the density of individuals in a habitat increases,
the suitability of the habitat will decrease until a second habitat becomes
more suitable and is selected. Eventually all habitats will have equal suitability.
This model assumes that all individuals are free to enter a habitat and will
select the habitat most suitable to them. In Virginia, the first level of
nest-site selection will occur at the landscape scale (Burger 1985) as
Gull-billed Terns, Common Terns, Black Skimmers, and American Oystercatchers
select between nesting on barrier island beaches, shellpiles on salt marsh
islands, or on wrack or Spartina on the marsh surface. The second level
of nest-site selection involves choosing a certain shellpile, the third
choosing a territory, and the fourth selecting a nest site within it (Burger
1985).
Nest-site selection is of
primary importance for the reproductive success of a breeding pair. The
selection of a nest site is influenced by physical factors, including
elevation, substrate, exposure, slope and aspect, and biological factors,
including presence of conspecifics and heterospecifics, vulnerability to
predation, and previous experience (Buckley and Buckley 1980). These factors
combine to define the nesting habitat chosen by a bird.
The elevation of a nest site
is particularly important on shellpiles that are subject to flooding. American
Oystercatchers have been shown to prefer higher nest sites (Lauro and Burger
1989), as have Gull-billed Terns (Clapp et al. 1983). American Oystercatchers
also used elevated platforms for nesting in Virginia (Nol and Humphrey 1994).
In an experiment using artificially constructed wrack-mats, both Common Terns
and Black Skimmers preferred higher elevation mats (Burger and Gochfeld 1990).
Because flooding is a major cause of nest failure among waterbirds nesting on
shellpiles in Virginia (Eyler et al. 1999), examining nest-site selection with
regard to elevation will be an important management component of research. I
expect that since higher nest sites will provide greater protection against
flooding and nest failure, species will select higher nest sites than are
randomly available on the shellpile. Since early nesters have first choice of
quality nest sites, it is expected they will choose sites of higher elevations.
For example, since American Oystercatchers arrive earliest in the spring, they
should chose the highest sites.
Choice of substrate on the shellpiles is limited to dense shell or tidally-deposited wrack, or a combination of the two. Nests of Gull-billed Terns (Parnell et al. 1995), Common Terns (Burger and Lesser 1978, Burger and Lesser 1979), Black Skimmers (Gochfeld and Burger 1994), and American Oystercatchers (Nol and Humphrey 1994) have all been found on wrack and shell. Nests on wrack can provide protection from flooding by acting as a floating raft, unless wind-driven waves break it apart (Burger and Lesser 1978, Lauro and Burger 1989, Gochfeld and Burger 1994). Substrate can also provide camouflage for eggs and chicks (Burger and Lesser 1978, Kotliar and Burger 1986, Mallach and Leberg 1998). Exposure of a nest site to sun, wind, and waves will also affect nest-site selection. The presence of vegetation near a nest site can affect exposure to sun and provide cover for adults, eggs, and chicks. Once chicks are old enough to move around, proximity to shade and shelter may improve chances of survival, and thus the area surrounding a nest may be an important factor in nest-site selection. However, thick vegetative cover can reduce visibility (Leberg et al. 1995). Black Skimmers had less vegetative cover than Common Terns around their nests in New York (Gochfeld 1978) and preferred areas with less than 30% cover (Gochfeld and Burger 1994). Gull-billed Terns prefer nest sites with sparse vegetation that are near areas with denser vegetation (Sears 1978). Common Terns on beaches in New York nested in areas with 10-25% vegetation cover, and avoided overgrown areas (Burger and Gochfeld, 1991). The exposure of a shellpile slope to wind or storm-driven waves may also influence nest-site selection. Nests on slopes facing an open lagoon will have a greater chance of flooding than will nests on marsh-facing slopes. Changes in wind direction may also affect which side of the shellpile is flooded (Burger and Lesser 1979, Burger 1982). To enhance success, birds are be expected to choose nest sites that minimize exposure to wind and waves, and provide protection for eggs and chicks (either through substrate or vegetation).
In addition to physical
factors, social factors are often important in nest-site selection. Competition
for nest space in colonial-nesting species can be very intense (Buckley and
Buckley 1980), especially when nest sites are limited, such as on shellpiles.
Oystercatchers typically arrive on last year’s territory in late March/early
April (Nol and Humphrey 1994) and lay their first clutches during the first
week of April, with a peak in mid-April (Nol and Humphrey 1994). Gull-billed
Terns arrive in late April to early May (Byrd and Johnston 1991) with nest
building beginning 5 to 25 days afterwards and egg-laying in mid-May to early
June (Parnell et al. 1995). Common Terns also arrive in late April/early May,
begin establishing territories the first two weeks in May, and lay eggs around
May 15 (Erwin 1979), but usually slightly later than Gull-billed Terns (R.M.
Erwin, pers. obs.). Black Skimmers arrive from late April to early May (Erwin
1979), with peak egg-laying the third week of May (Erwin 1979). Black skimmers
are usually the last of these species to lay eggs (B.R. Truitt, TNC; R.M.
Erwin, pers. obs.), but may renest as late as August in Virginia (B.R. Truitt,
unpubl. data).
As the first species to
arrive and establish nests, American Oystercatchers have the broadest range of
nest sites available to them. Their choice of prime, high-elevation nest sites
may limit species arriving later in the season from using these sites.
Aggressive and territorial behavior from birds (especially large-bodied species)
with established territories will further affect nest-site selection of
later-arriving birds. Fretwell (1972) described an “ideal despotic
distribution,” in contrast to the earlier-mentioned ideal-free distribution, in
which aggressive behavior from an individual with an established territory
deters a later-arriving bird from settling. The suitability of this habitat
will now not only be density-dependent but also dependent on the social
dominance of already-settled heterospecific individuals (Fretwell 1972). The
sequence of arrival at shellpiles, and the behavior of the early-arriving
birds, may influence the final distribution of nesting birds, and overall
species nest-site quality (e.g. higher elevations). Common Terns are known to
be the most aggressive of the four species (Erwin 1979), and this may influence
the frequency, intensity, and outcome of their aggressive interactions. Species
size may also influence the outcomes of aggressive encounters (American
Oystercatcher, Black Skimmer, Gull-billed Tern, Common Tern, ranking largest to
smallest). In addition, aggression may be density-dependent.
Selection of a nest site
within the colony is also an important factor in nest-site selection.
Aggressive territorial behavior could also serve to space individuals within a
habitat (Fretwell 1972). Solitary American Oystercatcher nests were spaced an
average of 124 to 190 m apart, depending on the habitat (Nol and Humphrey
1994). Nearest-neighbor distances for Gull-billed Terns ranged from 2 to 92 m
in Denmark, with an average of 22.4 ± 14.3 m (Moller 1981) and 4 to 16.5 m in
North Carolina (Sears 1978). Distances between Gull-billed Tern and Common Tern
nests ranged from 5 to 10 m, and Gull-billed Tern-Black Skimmer nests from 2.7
to 8.5 m (Sears 1978). Burger and Lesser (1978) found that Common Terns nested
a mean distance of 0.8 to 4.8 m apart on New Jersey salt marshes, depending on
substrate, size of island, and colony space, while Erwin (1977b) found
inter-nest distances of 1.6 ± 0.49 m on Virginia barrier islands. Pius and
Leberg (1997) reported that Black Skimmers nest, on average, 1.68 m from other
Black Skimmers, and 1.53 m from Gull-billed Terns in Louisiana. Mean
nearest-neighbor distance for Black Skimmers in salt marsh colonies ranged from
1.0 to 5.3 m in New Jersey (Gochfeld and Burger 1994). These distances,
especially for Gull-billed Terns, are greater than those I observed on the
shellpiles in 2001, where nests were often less than 1.0 m apart in densely
packed clusters of 8-10 Gull-billed Tern nests. Small shellpile area with
limited nesting sites probably causes nest densities to be higher on shellpiles
than on other, larger sites.
Black Skimmers commonly nest
among Common Terns and Gull-billed Terns in Virginia (Erwin 1977b, Erwin 1979).
Most of the nest distribution within Common Tern-Black Skimmer colonies can be
attributed to differences in microhabitat preferences between the two species
(Erwin 1979). Common Terns nested near vegetation while Black Skimmers nested
in open sand patches on barrier islands (Erwin 1979). Skimmer nests are often
clustered within the colony of terns they are nesting with (Burger and Gochfeld
1990). On Man & Boy Island in 2001 some Common Tern nests were located
within the area occupied primarily by Gull-billed Terns, but most nests were
segregated by species (R. Rounds, pers. obs.). In addition, Black Skimmer nests
were interspersed among Common Tern and Gull-billed Tern nests on North Conjers
in 2001 (R. Rounds, pers. obs.).
Vulnerability to predation
may also influence nest-site selection. Black Skimmers will abandon colonies
after unsuccessful breeding seasons due to high predation rates (Burger 1982).
Predation on marsh islands is usually limited to avian predators such as
Herring (Larus argentatus), Greater Black-backed (L. marinus) and
Laughing Gulls (L. atricilla), American Oystercatchers, Ruddy Turnstones
(Arenaria interpres),
and Great-horned Owls (Bubo virginianus) because frequent flooding of
the low-lying islands deters mammalian predators (Burger and Lesser 1979).
Colony-site selection may be affected by the presence of predators, while
nest-site selection will be a function of choosing a “safe” site from predators
and other potentially aggressive nesting associates, whether at the edge of a
colony near vegetative cover, or in the center of the colony.
The distribution of nests on
shellpiles is likely a function of the interaction between physical and
biological factors. Competition for territories is both inter- and
intraspecific, with birds arriving first at the site having an advantage in
nest-site selection. Prime nest sites will be occupied first, with later nests
filling in the empty space. As the season progresses, nearest-neighbor
distances will decrease, as nest density increases (Burger and Lesser 1979),
and thus the spatial patterns of nests on the shellpile will change.
Hatching Success
Because of the high flooding
risk to shellpile nesting, the physical location of the nest on the shellpile
is expected to have a strong influence on nest success. First, the elevation of
a nest can have a direct effect on nest success. The higher the nest, the less
likely it will be inundated and destroyed during flooding. Second, the exposure
of the shellpile to tides and waves will affect nest success. In addition,
since date of nest initiation may indirectly affect nest elevation, it may have
an effect on nest success as well. Substrate and vegetation could also
influence hatching success by providing better cover from predators or the sun.
Biological and social factors
that are involved in nest-site selection can also directly influence hatching
success. The date of nest initiation, which may influence nest elevation, is
also a function of parental age and experience, food resources, weather and
tidal regimes (Nisbet 1977, Burger and Gochfeld 1991). Clutch size may also
reflect parental quality as it reflects the amount of energy invested in an egg
(Nisbet 1977, Nisbet 1978), as well as being an indicator of resource
availability (Burger and Gochfeld 1991). The spacing of nests within the colony
is a function of nest-site selection, and may also influence reproductive
success. Within a colony, centrally located nests may have increased nesting
success because they are buffered from predators by peripheral nests (Buckley
and Buckley 1980). Burger and Gochfeld (1990) found that centrally located
Black Skimmer nests in New Jersey had higher hatching success, were closer to
Common Tern nests, and had larger clutches than nests closer to edges. However,
the opposite was found for Common Terns in New Jersey, perhaps because nests at
the edge of a colony were closer to sheltering vegetation (Burger and Gochfeld
1990). Since the shellpiles in Virginia are usually linear, colonies tend to
have more edge and little center.
Ground-nesting birds are
vulnerable to predation and therefore nest spacing and presence of concealing
vegetation are important factors influencing reproductive success (Buckley and
Buckley 1980). Laughing Gulls nest in close proximity to the shellpile colonies
at Wire Narrows and have been known to prey upon Common Tern chick and eggs
(Burger and Gochfeld 1991). At one of my colony sites, Ruddy Turnstones were
observed preying upon Gull-billed Terns in 1995 during several visits (R.M.
Erwin, pers. observ.) and are well-known egg predators that have predated tern
and skimmer colonies worldwide (Crossin and Huber 1970, Parkes et al 1971,
Loftin and Sutton 1979, Farraway et al 1986, Burger and Gochfeld 1990).
Predation rates also can vary throughout the breeding season, affecting early
and late nests differently (Shealer and Kress 1991). Avian predators are
expected to cause some nest failure, and the nest-initiation date and spatial
location of the nest will influence its probability of predation.
Sea Level Rise
Sea
levels along the Atlantic Coast of the United States are currently predicted to
rise 46 cm by 2100 (IPCC 1996). This could have devastating effects on the
amount of habitat available to nesting birds that is safe from flooding and
also on area of habitat available to nesting colonies. Flood tides are likely
to destroy some nests each year; however, the extent and timing of the flood
damage will be different from season to season depending on wind and storm
conditions. As sea levels rise, it can be expected that the number of floods
will increase each breeding season, along with the number of nests lost during
each flood tide. Tide data from Wachapreague, Virginia from 1980 to 2001
suggest a significant increase in frequency of flooding of marshes during the
May-July period (R.M. Erwin, unpubl. data).
Research Questions
To make the presentation
more organized, I have divided the series of questions into 2 major areas:
nest-site selection and hatching success.
Nest-site Selection
1.
How
does elevation affect nest-site selection?
a.
Do
species nest on experimental plots at a higher density than on control plots?
b.
Do
species select higher nest sites than what is generally available on the
shellpile?
c.
Are
there differences in the elevations of early versus late season nests?
2.
How
do substrate and exposure affect nest-site selection?
a.
Which
substrate(s) does each species prefer for nesting?
b.
What
level of exposure to open water does each species prefer at a nest site?
c.
Does
the choice of substrate and exposure differ from what is generally available on
the shellpile?
d.
Does
the choice of substrate and exposure differ between early- and late-season
nests?
e.
Do
substrates and exposure levels differ in elevation?
3.
How
does the presence of vegetation influence each species nest-site selection?
4.
How
does distance to the nearest-neighbor influence nest-site selection?
a.
Are
nearest-neighbor distances different between conspecifics and heterospecifics?
b.
Does
nearest-neighbor distance vary between early- and late-season nests?
c.
How
densely does each species nest?
d.
Does
the density of nests change between early- and late-season nests?
5.
Does
behavioral aggression during nest-site selection affect the number of nests on
experimental plots?
Hatching Success
6.
How
does elevation affect hatching success?
a.
Do
successful (hatched) nests have higher elevations than unsuccessful nests?
b.
Does
elevation affect hatching success in both early- and late-season nests?
c.
Does
flooding frequency vary between early- and late-season nests?
7.
Do
substrate and exposure affect hatching success?
8.
Does
presence of vegetation near a nest affect hatching success?
9.
Does
date of nest initiation affect hatching success?
10.
Does
clutch size vary between early- and late-season nests?
11.
Does
clutch size affect hatching success?
12.
How
does predation affect hatching success?
a.
Are
there differences in predation rates among shellpiles?
b.
Are
there differences in predation rates between early- and late-season nests?
c.
Does
nearest-neighbor distance affect predation rates?
13.
Does
nearest-neighbor distance affect hatching success?
14.
Do
biological or physical factors explain the most variation in hatching success?
Methods
Experimental Elevation of Plots (Question 1a)
From
March 11-14, 2002, before the focal species arrived at the shellpiles to nest,
plots on the shellpiles were experimentally elevated. We created 6 sets of
paired plots (one elevated, one control) at 4 of the shellpiles, and 4 paired
plots at the smaller South Conjers shellpile. The location of the paired plots
was randomly determined by laying a meter tape down along the long axis of the
shellpile, and using a random number tape to determine the distance to the
first plot of each pair. The status of this plot (elevated or control) was also
determined randomly. Each elevated plot was contoured similar to its paired
control plot, including addition of wrack where necessary. We attempted to make
experimental plots 15-20 cm higher than control plots. Fine shell was added to
the tops of elevated plots to mimic the natural shellpile features. Each plot was
2 m by 2 m. Based on previous experience at the shellpiles, we believed this
area was sufficient for at least 4 pairs of terns or skimmers to establish
nests. Control and elevated plots were at least 1 m apart, and there was at
least 1 m between each pair of plots. Stakes were put into the shellpile at the
four corners of each plot. All experimental plots were elevated using oyster
shell from lower or out-lying areas of the shellpile not used by nesting birds.
At Wire Narrows West two plots were partly elevated by laying a layer of wrack
at the bottom and covering it with shell. A third plot was elevated using a
pallet and covering it with shell. I measured the elevations of control and
elevated plots using a laser level. If the plot was located along a ridge,
multiple points were taken at different elevations. The increase in elevation
on experimental plots was not high enough to produce changes in predation
rates, or influence microclimate or other factors that might affect nest-site
selection.
I
set up six paired plots at Wire Narrows West. Three plots were located in an
area used by nesting Gull-billed and Common Terns in 2001 and three were
located in an area that had fewer, or no, nests in 2001. I also set up six
paired plots at Wire Narrows East. Three paired-plots were in areas high nest
density in 2001, and three in areas of low nest density. I created six paired
plots at North Conjers; however, lack of knowledge of previous nesting patterns
precluded identifying areas as used or unused. Only 4 paired plots were
established on South Conjers due to the small size of the shellpile, and no
areas of the shellpile were established as previously used or unused. I also
created six paired plots at Man & Boy. Three of the paired plots were in an
area of high nest density in 2001, and three in areas with fewer, or no, nests.
Random Points (Questions 1b, 2a, 2b)
I
located twenty-five random points on each shellpile and elevation, substrate,
slope and exposure were recorded for each point. Random points were located by
laying a measuring tape down the long axis of the shellpile and determining
distance to the point from a random number table. A second random distance
(0-10 m) was selected to go either left or right to locate the point. I
measured the elevation of random points using a laser level. Substrate was
limited to oyster shell, wrack, or a combination of the two. Slope and exposure
were recorded using an index combining the two variables (Table 1).
Exposure was based on exposure to waves. Areas of
the shellpile facing an expanse of marsh were “protected,” those facing open
water were “exposed,” while those on ridges or in the middle of the shellpile
were “neutral.” In addition, if a random point was located in a plot this was
recorded, along with whether the point fell within an area of high, low, or no
nesting density from 2001.

Nest Monitoring (all questions)
I
monitored the shellpiles twice a week for nesting activity from May 1 to July
31. To minimize disturbance to nesting birds, I visited the shellpiles for less
than 30 minutes, and to the extent possible, visited during the cooler hours of
early morning or evening. After clutches were initiated, I marked nests with 25
cm nails hammered into the shellpile within 6-8 cm of the nest scrape. All nests
on the shellpiles were monitored. I tied a metal tag with nest number and
species to the nail. I recorded the following during each visit: species,
content of the nest, substrate, slope/exposure, and presence of nearby
vegetation when the nest was first found. The status of each nest was assessed
during each subsequent visit. If a nest scrape was empty, I tried to determine
whether the nest had hatched or failed. Empty nests were examined for evidence
of flooding or predation. All nests with eggs that disappeared before hatching
without any evidence of nest fate (e.g. flooding) were assumed predated. Eggs
were also routinely felt for temperature and signs of abandonment. I assigned
new nest numbers for renesting attempts in a previously used scrape. I monitored
nests only until hatching, because determining fledging success is logistically
difficult without using enclosures (Eyler et al. 1999).
I
recorded the date of nest initiation (first egg laid) for each nest. If the
exact date was not known, an approximate date was determined from the date of
later-laid eggs or by back-calculating from the hatch date (incubation length
averages 22 days in Gull-billed Terns [Parnell et al. 1995], 21 days in Common
Terns [Parnell and Soots, 1979], 23 in Black Skimmers [Gochfeld and Burger
1994], and 26 in American Oystercatchers [Nol and Humphrey 1994]). If a nest
was found with a full clutch and subsequently failed, I estimated the dates of
nest initiation from the dates of previous nest visits. At the end of the
season, the elevation of every nest was recorded with a theodolite total
station unit.
Tide Gauges
I
set up temporary tide gauges at each shellpile to determine heights of spring
high-tides and storm waves. A piece of 120 cm long, 5 cm wide PVC with holes
drilled in the sides was inserted into the marsh near the shellpile. I added
ground cork to the PVC and the level of the cork after high tides was recorded
within 1-2 days of high-water events. In addition, I recorded the elevation of
each tide gauge using a theodolite total station unit.
Behavioral Observations
To
evaluate the role of social interactions, I observed bird interactions at Wire
Narrows West and East during the early stages of the breeding season from a
platform located 150 meters from Wire Narrows West and 190 meters from Wire
Narrows East. The presence of an observer on the platform at this distance did
not disturb the birds. I used a spotting scope, binoculars, and a cassette
recorder for observations. I began
observations of the shellpiles on May 1, 2002 and continued until May 29, 2002.
I discontinued observations when the majority of birds had established nests
and territories.
I
used focal-animal sampling (Altmann, 1974) for the behavioral observations. I
randomly selected an individual bird and observed it for as long as the bird
was visible, for a maximum of 10 minutes per sample. I discarded all
observations of less than 1 minute. I recorded all aggressive behaviors
observed during this period involving the focal animal, including behaviors
initiated and received by the focal bird. Aggression was ranked on 5 levels
ranging from no aggression to aerial stoop attack. I recorded the species of
each bird involved in an aggressive interaction, along with the approximate
stage of nesting of the majority of individuals of each species (pre-egg
laying, egg laying, incubating, hatch) and the outcome of the interaction. At
the beginning and end of each sampling session, a census of all the birds
within 2 and 5 m of the focal animal was conducted. If the focal animal was
located within either an experimental or control plot then the number of birds
within the plot and within 5 m of the focal animal was recorded.
Due
to the low nesting density in areas visible from the platform on Wire Narrows
West and East, I also used focal-area sampling (Altmann 1974). An area of the
shellpile about 10 m in length was observed continuously for 30 minutes. At
ten-minute intervals I recorded all birds in the observation area. During this
time all aggressive interactions between species were recorded as described
above.
Data Analysis Methods
Statistical
I conducted a power analysis (O’Brien 1998) using UnifyPow and SAS to determine if 25 random points were enough for t-test comparisons between random point elevation and nest elevation. I used chi-square analysis on contingency tables to test for many of the analyses in order to determine if observed frequencies matched expected frequencies. I used contingency tables for all analyses of nest-site selection and hatching success involving substrate, exposure, and vegetation. I also used contingency tables to analyze data involving hatching success and nest-initiation date, flooding, and predation. Contingency tables were used to test for goodness-of-fit for nest-site selection data and independence for hatching success data. In order to test for differences between means, T-test and Analysis of Variance (ANOVA) were used to analyze some of the elevation, clutch-size, and nearest-neighbor data. Because I was interested in what factors explained hatching success, I used logistic regressions to examine if elevation, clutch size, lay date, and nearest-neighbor distance influenced hatching success. All data analysis was performed using the SAS statistical package (SAS 1999) except nearest-neighbor distances which were calculated using the Distances function in SPSS (SPSS 2002).
Description of Variables used in Analysis
Substrate: Substrate was divided into shell, wrack (washed up
dead vegetation) and shell-wrack. Shell-wrack indicates that nest substrate was
a combination of shell and wrack.
Slope-Exposure Index: Because of small sample
sizes, I combined the flat and slope categories into exposed, protected, and
neutral. Observation in the field made it clear that exposure to flooding was a
more important variable to nest success than flat or sloped. In addition, the
small number of nests on protected sites made analysis difficult so these nests
were combined with either neutral or exposed sites depending individually on
their location.
Vegetation: I recorded “vegetated” nests as being those within
30 cm of a patch of vegetation of any type, regardless of density.
Clutch size: The clutch size for each nest used in the analysis
was total number of eggs in each nest. Nests that failed before full clutch
size was reached were excluded from the analysis.
Lay Date: Lay date was determined from the hatching date and
the date the nest was first observed. May 1 was chosen as day 1 since all
nests, (with the exception of American Oystercatchers), were initiated after
this date.
Early vs. Late nests: All nests initiated after
June 7 were considered “late” nests. June 7-12 was the week of a large early
season flood that destroyed 47% of all nests. Soon after this flooding, a new
wave of nest-initiation began of “late” nests.
Nearest-neighbor: The shortest distance from the focal nest
to a neighboring nest. Because some nest-stakes were lost in floods, the
location of each nest could not be determined. These nests were left out of all
nearest-neighbor analysis, and so the nearest-neighbor distance may be slightly
overestimated.
Areas: The area of “used habitat” was calculated in
ArcView by tracing the outer edges of the colony, or focal nests. The more
inclusive “available habitat” is the area of the whole shellpile, including
areas used by nesting birds. I calculated nesting density using both “used” and
“available” areas.
Nest Outcome: I grouped nests as either “hatched” (≥ 1
egg) or “not hatched” for analysis. If one egg of the clutch hatched then the
nest was considered successful for analysis purposes. If I found a dead chick
on the next visit, this nest was still classified as “hatched.” I considered
nests whose eggs survived more than 20 days to be successful, even if their
eggs or chicks were not found on the next visit. Most nest predation occurs in
the first 2 weeks of nesting (Erwin and Smith 1985, Eyler et al. 1999). Some of
these nests also showed signs of chick presence (fecal matter, flattened wrack)
indicating hatching. Nests whose outcome could not be determined were listed as
unknown, and excluded from analysis.
Predation: I considered a nest predated if evidence of
predation (broken eggs, yolk in nest) was found at the nest scrape or in the
nearby vicinity. In addition, eggs that disappeared before they should have
hatched were considered predated.
Flooding: Because a new wrack line is deposited after a
flood, determining nest flooding was unequivocal. The new wrack usually
contained many of the eggs that were washed out of their scrapes. Indications
of flooding also included deposition of new shell over the nest scrape, and
flattening of the scrape by wave action. I relocated many nests by digging
through shell and wrack until the nest stake was located.
Abandoned nests: Nests were considered abandoned when eggs
were felt to be cold to the touch, or when they were present more than 5 days
beyond the expected hatch date (30 days for Common Terns).
GIS
I
used ArcView 3.2 (ESRI 2002) and ArcInfo (ESRI 2002) to analyze Global
Positioning System (GPS) data collected from the GPS survey units.
Tides
The
tide data used for this research was provided by the Virginia Institute of
Marine Science (VIMS 2002) and NOAA websites (NOAA 2002) for Wachapreague,
Virginia.
Results
Nest-Site Selection
Physical Factors
Experimental Plots (Question 1a)
South Conjers had the
highest percentage of nests on elevated plots (8%) compared to just 2% at Wire
Narrows East (Table 2). In addition, 10% of late nests at Wire Narrows West
were on elevated plots. At all five shellpiles, nesting birds did not select
experimental plots as nest sites at different frequencies than compared to
controls (Table 2). The density of nests on experimental plots was lower than
on the used area of the shellpile as a whole except at Wire Narrows West. At
Wire Narrows West the density of nests on experimental plots was 2.38 nests/m2
while the overall density on the used area of the shellpile was 0.9 nests/m2.
Elevation (Question 1b&c)
The power analysis found that with a sample size of 25 random points and 50 nests, a standard deviation for elevation of 0.122 m (based on the random point standard deviation), and a difference in elevation of 0.1 m, the power of the test was over 0.9. Because sample sizes were larger, and standard deviations lower, than used in the power analysis, the power to detect differences between random points and nests elevations was high.
There were shellpile and
species differences in nest elevations (Figures 2-3, Table 3, Appendix 3).
Common Tern nests at both Wire Narrows shellpiles were significantly lower than
random points, while at the two Conjers shellpiles Common Tern nests were
significantly higher than random points. At 3 shellpiles late Common Tern nests
were significantly higher than early nests (Figure 3). Gull-billed Tern nests
were significantly higher than random points at one colony, and lower at
another colony, although not significantly. Black Skimmer nests were not
different in elevation than random points. American Oystercatcher nests were,
on average, higher than random points at all sites, although not significantly
(Figure 2).
Substrate and Exposure (Question 2)
Species
differed in their utilization of substrate and exposure of nest to open water,
and site differences existed as well (Figure 4, Table 4-5). At all sites, nest
substrates (Table 4) and exposures (Table 5) were chosen differently from that
available. Overall, Common Terns preferred wrack on exposed slopes (39% of all
nests), Gull-billed Terns and Black Skimmers selected shell on neutral slopes
(62% and 61% of all nests, respectively), and American Oystercatchers preferred
shell on neutral slopes (58% of all nests). At Wire Narrows East and South Conjers,
the number of Common Tern nests on wrack decreased over the season (Appendix
6). At Wire Narrows East and Man & Boy the number of Common Tern nests on
exposed slopes decreased, and on neutral slopes increased, between early and
late nests (Appendix 7). For all shellpiles except North Conjers, the elevation
of neutral nests was higher than that of nests on exposed slopes (Table 6), and
at 3 shellpiles, shell nests were significantly higher than nests on either
other substrate (Table 7).
Vegetation (Question 3)
No
vegetation was present on North Conjers or Man & Boy shellpiles. Only
Gull-billed Terns at Wire Narrows West nested near vegetation at a high
frequency (70% of nests, Table 8). Gull-billed Tern and Common Tern nests near
vegetation were significantly lower than were nests remote from vegetation
(Table 8).
Biological
Factors
Nearest-Neighbor Distance (Question 4a&b)
The
average distance from each species nest to its nearest-neighbor illustrates
major species and site differences (Table 9). The average distance from each
species to its nearest-neighbor of each species is shown in detail in Appendix
8. In assessing seasonal effects (question 4b), late Common Tern nests were
closer together than early nests at all sites but Man & Boy (where nests
were the same distance apart, Table 9). Both Black Skimmer and Gull-billed Tern
late nests were closer to nearest-neighbors than early nests.
Density and Area (Question 4c&d)
The
area of each shellpile and the density of nests in both used and available
habitats are shown in Table 10. The density of Common Tern nests in used areas
ranged from 0.47 nests/m2 at North Conjers to 0.9 nests/m2
at South Conjers, and densities of nests also varied between early and late
nests (Table 11). Gull-billed Tern nest density ranged from 0.7 nests/m2
to 1.9 nests/m2 and also varied between early and late nests (Table
11).
Behavioral Observations (Question 5)
A total of 30 hours of
behavioral observations were conducted. The observations were limited since
most of the early-nesting terns did not establish nests in areas visible from
the observation platform. While all 6 plots on Wire Narrows West were visible,
and 3 plots on Wire Narrows East were visible, very few birds nested on these
plots, thus the number of focal animals was limited. Further, nesting density
was low in most of these areas and that may have limited interspecific
aggressive interactions. Once focal area sampling was used, more birds were
included in the observations; however due to the limited area of the visible
shellpile used by nesting birds, repeated observations probably were made of
the same nesting pairs (although, without marked birds, this is speculative).
However, qualitative data were collected that shed some light on behavioral
interactions during nest-site selection, and how these processes may have
affected nest-site selection on experimental plots.
The
number of interspecific aggressive interactions I observed illustrated species
asymmetries (Table 12). All six of the occurrences of American Oystercatchers
chasing Black Skimmers occurred at the time of hatching of the focal American
Oystercatcher’s nest. The Black Skimmers were all within 10 m of the nest, and
the American Oystercatcher pair repeatedly chased the birds from the area. Four
days later, on my next visit to the shellpile, many Common Tern, Gull-billed
Tern, and Black Skimmer nests were found in the area of the Oystercatcher nest.
In fact a Common Tern pair was using the same scrape as the Oystercatcher nest.
The aggressive behavior of the American Oystercatchers, therefore, appeared to
have no lasting impact on nest-site selection of the other species using the
shellpile.
Seventeen
of the aggressive interactions initiated by Gull-billed Terns towards Black
Skimmers were from a pair of Gull-billed Terns nesting on experimental plot 3E
on Wire Narrows West. The aggressive behaviors were first observed on May 25
and eggs were first found in this nest on May 27. Thirteen of the aggressive
interactions resulted in Black Skimmers leaving the area (Table 12). However,
despite the territorial aggression exhibited by the Gull-billed Tern pair, on
June 4 a Black Skimmer nest was found on plot 3E in the same place the Black
Skimmer pair had occupied earlier. The Gull-billed Tern nest was still active
at this time. By June 18 there were 3 Gull-billed Tern and 3 Black Skimmer
nests on plot 3E. The aggressive behavior of the first Gull-billed Tern nest
initiated on plot 3E did not affect future use of the plot by pairs of either
species.
Common
Tern nests on Wire Narrows East and West were primarily found in low-lying
vegetated areas and were not visible from the observation platform. However,
one area used by both Gull-billed Terns and Common Terns was visible. Very few
aggressive interactions between the two species were observed (Table 12) and
both species nested in close proximity in this area.
Hatching Success
The
summary of hatching success indicates some species differences (Figure 5 and
Appendices 9-13).
Physical
Factors
Elevation (Question 6)
Elevation
had a significant positive effect on hatching success for Common Terns at all
shellpiles except North Conjers (Table 13). At 3 shellpiles, however, hatching
success was improved only in early nests (Table 14). Hatching success improved
with decreasing elevation for late season Gull-billed Terns at Wire Narrows
West (Table 14). Hatching success of Black Skimmers was not affected by nest
elevation.
For
all shellpiles, except North Conjers, significantly more early nests flooded
than late nests because of the high-water period of June 7, 2002 (Figure 6).
For all the shellpiles (excluding North Conjers) there was a significant
difference (P<0.001) between the elevations of flooded nests and non-flooded
failed nests. The elevations of failed nests that did not flood were not
significantly different from the elevations of nests that hatched. This
indicates that elevation only affects nest success with respect to flooding
frequency and does not affect other factors that might influence a nests’
outcome.
Substrate and Exposure (Question 7)
There
was no significant difference in hatching success between nests on shell,
wrack, or shell-wrack at Wire Narrows West, South Conjers, or North Conjers
(Table 15). At Wire Narrows East and
Man & Boy hatching success was highest on shell-wrack and lowest on wrack.
All sites yielded higher hatching success for Common Terns and Gull-billed
Terns on neutral sites than exposed sites (Table 16).
Vegetation (Question 8)
Hatching
success is significantly lower at nest-sites near vegetation for Common Terns
at both Wire Narrows sites, but higher for Gull-billed Terns at Wire Narrows
West (Table 17).
Biological Factors
Nest-Initiation Date (Question 9)
Peak nest-initiation dates
for each species are shown in Figures 7-9. Common Terns had higher hatching
success later in the season at all sites (Table 18). There were no seasonal
differences in Gull-billed Terns or Black Skimmer hatching success.
Clutch Size (Question 10&11)
Average
clutch size for early and late nests did not show consistent seasonal declines
(Table 19). Black Skimmers had significantly smaller clutch sizes in later
nests; I found 4 and 5-egg clutches early in the summer, but none late. Common
Terns at Wire Narrows East, Man & Boy, and South Conjers also had
significantly smaller clutch sizes in later nests.
The effect of clutch size on
hatching success revealed that, for all species, as clutch size increased,
hatching success increased (Fig. 10). At four sites, Common Tern nests with
2-egg clutches had the highest hatching success.
Predation (Question 12)
At all five shellpiles there were no statistically significant differences in the frequency of predation between early- and late-season nests (Table 20). However, variation in predation rates among shellpiles was found in the early season primarily because 62% of early-season nests at North Conjers were predated (Table 20). The relationship between predation and nearest-neighbor distance showed that only for Common Terns at Wire Narrows West were denser nests more immune from predation than were more dispersed nests (Table 21).
Ruddy
Turnstones had a substantial impact on nest success in very early-season nests
(Table 20). On May 18, 13 Gull-billed Tern
nests were found on Wire Narrows East, five in an area visible from the
observation platform. On May 19, a Ruddy Turnstone was observed predating nests on Wire Narrows East.
Only two Gull-billed Terns were observed incubating on the morning of May 19. A
Ruddy Turnstone was observed walking from one nest to the next, eating the
contents. A visit to Wire Narrows East found that nine Gull-billed Tern nests
had been predated. In total, Ruddy Turnstones predated 12 of the first 14
Gull-billed Tern nests on Wire Narrows East. Ruddy Turnstones were observed at
the colonies from May 9 through June 21. Ruddy Turnstones reappeared in late
July when the majority of nests were hatched, but no nests were predated by
turnstones in the late season.
Nearest-Neighbor Distance (Question 13)
Nearest-neighbor
distance significantly affected hatching success for Common Terns at Wire
Narrows West, East, and Man & Boy (Table 22). Black Skimmer hatched nests
were closer together than were failed nests, although the results were not
significant. Gull-billed Terns had similar nearest-neighbor distances for
hatched and failed nests.
Flooding
There was
one major episode of flooding beginning on June 7 and continuing through to
June 14. During this time period, 47% of all the nests on all shellpiles were
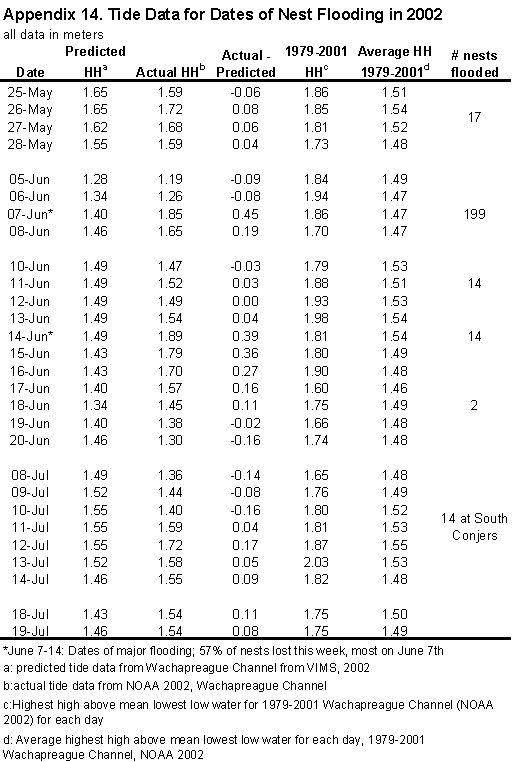
flooded. On June 7, the actual tide in Wachapreague Channel (NOAA 2002) was
0.45 m higher than the predicted tide (VIMS 2002, Appendix 14). These high
tides may have been caused by a low-pressure system offshore. Although most
nests were flooded on June 7, a few more were lost later in the week. On June
14, the highest high reading was 1.89 meters above mean lowest low water, and
the highest reached all summer 2002. This value was the highest reading since
1979 for that date. This flood had major effects on all 5 sites. Wire Narrows
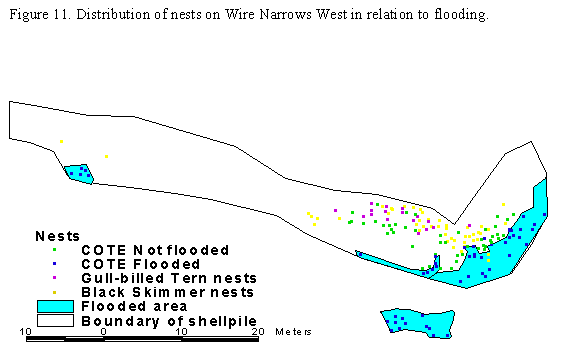
West had the largest amount of used habitat flooded (Figure 11, Table 23),
while Man & Boy had the smallest area flooded (Table 23). The number of
nests in the flooded areas decreased from early to late nests (Table 23) at all
shellpiles except Wire Narrows West.
Some
nests were also lost to flooding during the late May spring high tides.
However, these tides were barely above predicted (0.05 m) and did not reach
previous highest high records and were below the average of the previous 20
years. Floods also threatened nests in late July, but these tides did not reach
the extent of early June.
Overall Hatching Success (Question 14)
Gull-billed
Terns were the only species that had more nests hatch (Figure 5, Figure 12)
than fail, all at Wire Narrows West. American Oystercatchers had the lowest
nest success, with a conservative estimate of 26% of nests hatching, although
the fate of 32% of nests was unknown. When I used a multiple logistic
regression to determine which variables (elevation, date of nest-initiation,
clutch size or nearest-neighbor distance) were most important in predicting
hatching success, the results differed among sites and species (Table 24).
Elevation and date of nest-initiation were important in determining hatching
success for Common Terns at all sites analyzed (n=3), with higher elevation and
later date of nest-initiation improving hatching success.
Figure 2. Nest elevation (mean ± 1
SD) vs. random point (RP)
elevation at 5 shellpiles for Common Terns (COTE), Gull-billed Terns
(GBTE), Black Skimmers (BLSK), and American Oystercatchers
(AMOY).Shellpile codes: WNW = Wire Narrows West, WNE = Wire Narrows East,
MB = Man & Boy, SC = South Conjers, NC = North Conjers. *Indicates significant difference from random point
elevations: WNW-COTE T=-4.75, P<0.001, COTE nest elevations lower than
random points; WNW-GBTE T=2.86, P=.008, nest elevations higher than random
points; WNE-COTE T=-4.76, P<0.001, nest elevations lower than random
points; SC-COTE T=2.51, P=0.01, nest elevations higher than random points;
NC-COTE T=6.53, P<0.001, nest elevations higher than random points. Figure 3. Common Tern nest elevation (mean ± 1 SD) at
3 shellpiles (see Fig. 2 for site codes): early vs. late nests. * Significant
difference between early nest elevation and late nest elevation. Early
nests are significantly lower than late nests. WNW: T=-2.93, P=0.004,
n=187 nests; WNE: T=-10.34, P<0.001, n=139 nests; MB: T=10.34,
P<0.001, n=153 nests. Figure 4. Substrate and exposure use
by 4 waterbird species nesting at 5 shellpiles sites, data combined. Figure 6. Flooding in early and late
nesting seasons at 5 sites. *Results from chi-square analysis on contingency
table. WNW χ2=52.15, P<0.001, n=309; WNE χ2=89.77,
P<0.001, n=177; MB χ2=52.96, P<0.001, n=178; SC χ2=14.96,
P<0.001, n=99. There was no significant difference for NC. Figure 10. Hatching success versus clutch size in 5
species of waterbirds. Results of logistic regression analyzing effects of
clutch size on hatching success. WNW-BLSK: χ2=9.76, P=0.02, n=52;
WNW-COTE: χ2=4, P=0.045, n=167; WNW-GBTE: χ2=5.26, P=0.02, n=47;
MB-COTE: χ2=8.33, P=0.004, n=130. Hatching success increases with
larger clutch sizes. Figure 12. Hatching success of Common
Terns, Gull-billed Terns, and Black Skimmers at each shellpile.































Discussion
Nest-Site
Selection
Physical Factors
Experimentally elevated plots were not preferentially selected for at any of the shellpiles, or by any species, despite the extra protection from floods elevated plots provided to nesting birds. A number of other factors involved in nest-site selection could explain this including substrate, vegetation and previous experience at nesting sites. Common Tern nests were only significantly higher than random points at North and South Conjers, which account for only 20% of all Common Tern nests on the shellpiles. Eighty percent of Common Tern nests, therefore, were either of similar elevation to random points or lower. Because Common Terns did not select nest sites based on high elevation locations, elevated plots were not selected for preferentially. In addition, the combination of choosing to nest on wrack and near vegetation may have contributed to the small number of Common Tern nests on elevated plots at both Wire Narrows shellpiles. Gull-billed Tern nests at Wire Narrows West were higher than randomly available, and were concentrated around one of the pairs of experimental plots. At Wire Narrows East and Man & Boy, Gull-billed Tern nests were not different in elevation from random points. Gull-billed Terns may have been primarily selecting sites based on previous nesting experience at these shellpiles. For example, nests at Man & Boy were at the same place as nests had been found in 2001, on a low section of one ridge.
Nine percent of Black
Skimmer nests were in elevated plots. Black Skimmers chose to nest on shell
preferentially, and their nest elevation was not different from what was
available. Black Skimmers choose to nest among Gull-billed Terns and Common
Terns (Erwin 1977b, Pius and Leberg 1997), and the location of earlier tern
nests may have influenced their nest-site selection. American Oystercatcher
nests were higher than random points at all sites, and two nests (11%) were in
elevated plots.
Although
elevation may not be the most critical factor in initial nest-site choice, it
may become more important later in the season. At Wire Narrows and Man &
Boy, late nests of Common Terns were significantly higher than early nests, and
late nests at Man & Boy were significantly higher than random points. The
flooding on June 7 may have caused birds to renest at higher elevations,
especially because for most of the week after June 7 areas of the shellpiles
that Common Terns had previously nested on were underwater.
With
respect to substrate, Common Terns at all shellpiles used wrack at a higher
frequency than that randomly available on the shellpile. Burger and Lesser
(1978) pointed out that wrack provides good camouflage for the dark brown
Common Tern eggs. This is especially true compared to the white of bleached
oyster shells. Burger and Lesser (1978) found that over 80% of Common Tern in
New Jersey on salt-marsh islands nested on wrack. Eighty-two percent of Common
Terns in this study used either wrack or shell and wrack as nest substrates.
The Common Terns nested on wrack that had been deposited during earlier winter
and spring storms, and also on wrack that was deposited during the June 7
flood. At Wire Narrows East, Man & Boy, and South Conjers, the percentage
of nests on wrack decreased during the season. This change may be a result of
birds moving to higher or different areas than what was flooded in the early
season, since wrack nests had the lowest elevations at those sites. However,
wrack nests were also lower at Wire Narrows West yet Common Terns selected
wrack at the same frequency in early and late nests. Many late Common Tern
nests at Wire Narrows West were on the wrack line deposited by the June 7 flood
and this may account for the difference.
The
choice of shell as a nest-site substrate for Gull-billed Terns (69%) and Black
Skimmers (75%) suggest that their lighter-colored eggs (relative to Common
Terns) may be better camouflaged on shellpiles. Leberg et al. (1995) also found
that Gull-billed Terns disproportionately selected shell for nesting, but felt
that Black Skimmer nest choice was more influenced by the presence of nesting
Gull-billed Terns than the actual substrate. Shell nests had the highest
elevations at all the shellpiles used by Gull-billed Terns and Black Skimmers
and this also may have contributed to their preference of shell as a substrate.
American Oystercatchers, another species with light, creamy colored eggs, used
shell for 95% of nests, which is typical of other locations for this species
(Nol and Humphrey 1994).
Exposed
sites were more attractive to Common Terns than randomly available sites at
Wire Narrows and South Conjers. At Man & Boy and North Conjers Common Terns
chose neutral sites preferentially. Overall, 62% of Common Tern nests were at
exposed sites. At Wire Narrows the exposed sites were primarily low-lying
areas, near the tide line, with sparse vegetation. Common Terns may have been
attracted to these areas because these are the areas where wrack had been
deposited and because of the presence of vegetation for protection and cover.
At Man & Boy and North Conjers there is little vegetation cover, and at
North Conjers there was a lot of wrack on neutral sites. At all shellpiles
exposed sites had lower elevations than neutral sites. Similarly, Burger and
Lesser (1978) found that most Common Tern colonies on marsh islands were on the
side of the island exposed to open water. Gull-billed Terns, Black Skimmers,
and American Oystercatchers all preferred neutral sites, and generally avoided
the low-lying vegetated areas that Common Terns preferred.
Vegetation
also may have influenced nest-site selection. Common Terns, Gull-billed Terns,
and Black Skimmers will nest near vegetation if present and Gull-billed Terns
particularly seemed to select sites near vegetation, if the vegetation was on
higher areas of the shellpile. Kotliar and Burger (1986) reported Common Terns
nesting near short, sparse vegetation that is similar to what was available on
the shellpiles. Erwin (1979) found that Common Terns nested near vegetation,
with Black Skimmers in more open areas among the terns, while Gochfeld (1978)
found that Black Skimmers had less cover at their nests than Common Terns. At
Wire Narrows West, the areas with vegetation that Gull-billed Terns and Black
Skimmers used were much more open, with one or two plants every few meters. The
areas with vegetation used by Common Terns tended to have a denser distribution
of plants, though most were still very short with open space in between plants.
The presence of vegetation near a nest to provide cover and protection appears
to be an important factor in nest-site selection. Areas of Wire Narrows West
with dense vegetation were devoid of nests, but chicks were seen hiding for
cover in these areas. Only two American Oystercatcher nests, both at Wire
Narrows West, were near vegetation. In addition, vegetation may have influenced
use of experimental plots. The elevated plot on Wire Narrows West with a high
density of nests had a plant growing in the middle of the plot. Many
Gull-billed Tern nests were around this plant (and similarly around another
plant at Wire Narrows East) and this may have contributed to their use of this
plot for nests.
Biological and Social Factors
Common
Tern nearest-neighbor distances are similar to those reported elsewhere (Erwin
1977a, Burger and Lesser 1978), although on the lower end. Burger and Lesser
(1978) suggested that nearest-neighbor distance varied as a function of habitat
area. This would explain why nearest-neighbor distances are low on the
shellpiles since the area used by nesting birds at the shellpiles was limited.
Gull-billed Terns nearest-neighbor distances are much closer than reported
elsewhere (Sears 1978, Moller 1981). However, distances between conspecifics
were farther (1.15 m to 3.4 m). Black Skimmer nearest-neighbor distances were
on the lower end of what is reported elsewhere (Gochfeld and Burger 1994, Pius
and Leberg 1997). American Oystercatcher nests were very close together for a
solitary species at South Conjers (7.83 m, n=5) although distances at the other
shellpiles were greater than 20 m. The highest American Oystercatcher nest
density Lauro et al. (1992) found in New York and Virginia was 1.3 and 0.1
nests/km2, respectively. American Oystercatcher nest density on
South Conjers was 48 nests/km2. Lauro et al. (1992) found that at
the highest nest densities in New York, American Oystercatchers were breeding
communally, possibly because of a shortage of high quality territory. There was
no evidence of communal nesting at any shellpiles despite the high density of
American Oystercatcher nests at South Conjers, North Conjers, and Wire Narrows
West. Nearest-neighbor distances
changed significantly between early and late nests for Common Terns and
Gull-billed Terns at Wire Narrows West, with later nests being closer together.
Burger and Lesser (1979) also found that birds renesting after a storm nested
closer together, probably because of a decrease in suitable habitat.
At
Wire Narrows West, Gull-billed Terns nested the closest to other nests. If
nearest neighbor distance serves as a proxy for territory size (Burger and
Gochfeld 1990) then Gull-billed Terns had the smallest territories, while
Common Terns had the largest. In contrast to this study, Erwin (1977a) found
that Black Skimmers were more widely dispersed than Common Terns. Black
Skimmers nested in close proximity to both Common Terns and Gull-billed Terns
at Wire Narrows West in the only area of bare shell used by nesting birds.
Because Black Skimmers chose to nest near Gull-billed Terns and Common Terns,
possibly to improve nest defense (Erwin 1977b, Pius and Leberg 1997), and also
preferred open sites on shell, their nest density might have been higher than
expected because of the small area of the used area of the shellpile that
suited their preferences.
The
highest nesting densities were reached at Man & Boy with 1.92 nests/m2
for Gull-billed Tern nests. These nests were in a small patch of the shellpile
that was also used by Gull-billed Terns in 2001; previous nesting experience at
the shellpile may explain the high density of nests in this area.
Aggressive interactions between species did not play
a significant role in nest-site selection, at least at the Wire Narrows sites I
observed. The locations on the shellpiles where aggressive interactions were
observed eventually had high numbers of nesting birds of all species. It is
unlikely that the small number of nests on elevated plots was due to the
aggressive defense of a nest territory by early-nesting birds.
In
addition to the factors mentioned above, the locations of American
Oystercatcher nests seemed to play a role in nest-site selection for Common and
Gull-billed Terns. American Oystercatcher nests were established in April on
all five shellpiles, in some cases probably before the terns and skimmers had
arrived. Subsequently, many early Common Tern and Gull-billed Tern nests were
located around the early Oystercatcher nests, or in some cases, in the
Oystercatcher scrape. In many cases these nest locations became centers of the
future colony. For example, at Wire Narrows West there were three American Oystercatcher
nests in May when the terns arrived. These three nest locations became the
center of the three major nesting areas of the colony. Early nests of terns
were established near or in American Oystercatcher nests, and these areas
continued to be used throughout the breeding season. This pattern is probably a
combination of social attraction influencing nest-site selection, along with
previous nesting experience at the shellpiles.
At Wire Narrows West and East, and Man & Boy, in addition to the unused elevated plots, extensive areas of shell at high elevations and closer to the marsh (and so protected from flooding) were not used by any nesting birds. At Wire Narrows some of these areas, because of their proximity to the marsh, were also close to nesting Laughing Gulls. The Laughing Gulls, during the pre-egg laying stage, often stood and defended the area of the shellpile adjacent to their nests. Since Laughing Gulls are known predators of tern and skimmer nests, the birds may avoid nesting on these areas of the shellpiles. The only two late Gull-billed Tern nests at Wire Narrows East were located in a high, protected area of the shellpile near Laughing Gull nests, and were predated. However, I was unable to determine the cause of the predation. However, no Laughing Gulls nested near the colonies at Man & Boy and yet many protected areas of the shellpile were not used.
Nest-Site Selection Summary
Contrary
to expected, Common Terns did not choose higher nest-sites than were randomly
available on shellpiles. While Common Terns did nest in high locations, the
majority of their nests put them at significant risk of flooding. Over 85% of
Common Terns nested with 2 m of their nearest-neighbor, indicating that
location of other nesting birds is a strong selective force. This can also be
seen by the congregation of early nests in areas with an American Oystercatcher
nest. The role of previous experience of use of the shellpiles cannot be
assessed since accurate locations of Common Tern nests in 2001 or earlier are
unknown.
Gull-billed Terns at Wire
Narrows West nested in an area protected from flooding and at higher
elevations. At Wire Narrows East most nests were predated before the June 7
flood; however, if those nests had been active some would have flooded. At Man
& Boy Gull-billed Terns nested on a low-lying area of the shellpile that
was completely flooded on June 7. This area was also used by nesting
Gull-billed Terns in 2001 and it is possible that this part of the shellpile
used to have a higher ridge and more protection from flooding. Since the
shellpiles are frequently changed in shape by winter storms, Gull-billed Terns
may initially have selected this part of the shellpile when it was higher and
now continue to use the same site. At all sites, Gull-billed Terns primarily
selected nest sites on shell and on neutral slopes, and in close proximity to
other nests.
Black Skimmers at Wire Narrows West primarily nested in the higher shell areas that were used by both Gull-billed Terns and Common Terns. They picked the area used by both tern species with shell as the primary substrate. Since Black Skimmers chose colony sites based on locations of Gull-billed and Common Tern colony sites (Erwin 1977b, Pius and Leberg 1997), their nest-site selection was probably strongly influenced by social stimulus of nesting near already established tern nests at the shellpiles.
American Oystercatcher
nest-site selection is probably based primarily on physical factors, since no
other species are present, or at least nesting, on the shellpiles when American
Oystercatchers begin nest-site selection. Previous experience nesting at the
shellpiles may also play a strong role (Nol and Humphrey 1994).
Marsh islands provide marginal habitat for nesting terns, skimmers, and oystercatchers because of their low elevations. Providing elevated sites for nests met with very limited success. The elevated plots did not provide the habitat preferred by Common Terns, and may not have been located in areas previously used by Gull-billed Terns, especially at Man & Boy where an elevated plot in the Gull-billed Tern nesting area probably would have protected the nests from flooding. Nest-site selection varied among species and shellpiles, making it difficult to predict what areas of a shellpile will be used or avoided by nesting birds. Colony and site-specific factors appear to be important when examining nest-site selection.
Hatching
Success
Physical Factors
For
all species at all shellpiles, the elevation of a nest influenced its hatching
success. Flooding of nests on marsh islands is generally higher than on barrier
island beaches (Burger and Gochfeld 1991), however the marsh islands are
protected from ground predators. Buckley and Buckley (1980) suggest that Common
Tern’s ability to adapt to salt marsh nesting may be related to their ability
to relay after losses to flooding. Late nests, many assumed to be renesters,
accounted for between 43-57% of all Common Tern nests (depending on the
shellpile) indicating that many birds renested after the June 7 floods.
Common
Tern nests with higher elevations had higher hatching success at all shellpiles
except North Conjers. The June 7 floods had a significant effect on these
results. At Wire Narrows and Man & Boy early nests had improved hatching
success at higher elevations. Black Skimmers hatching success was not affected
by elevation and Black Skimmers also had the lowest rates of flooding (Figure
5) of all species, since they primarily nested in an area protected from
floods. The hatching success of Common Terns at South Conjers was not affected
by elevation in the early season, but only in the late, a reverse trend than
the other shellpiles. Thirty percent of late nests at South Conjers flooded, a
much higher rate than any other shellpile (Figure 11), and this may have
affected the result. It is possible that because of the small size of the
shellpile, birds were forced to renest in low-lying areas that flooded early
because of competition for limited nesting sites.
Because
of the unpredictable nature of flooding, the date of nest-initiation will be
important to hatching success, even if the date is not consistent across
different years. Nisbet (1977) found that younger Common Terns nested later,
and that younger birds had nests with lower elevations (Nisbet et al. 1984). In
addition, late nests also often have lower reproductive success than early
nests (Spendelow 1982, Nisbet et al.1984, Burger and Gochfeld 1990). In this
study, however, late Common Tern nests were higher than early nests and no
major floods occurred during the late season, therefore hatching success
improved.
Substrate
seemed to affect hatching success. Hatching success at Man & Boy, Wire
Narrows East and South Conjers was lowest for nests on wrack. Since elevations
of wrack nests were lower than that of other substrates, the increased chance
of flooding for a wrack nest probably made hatching success lower. At Wire
Narrows West and North Conjers, areas with wrack occurred on high grounds and
were protected from flooding, therefore hatching success was not different
between the different substrates. Nests that were exposed to open water had
significantly lower hatching success than exposed sites because of their lower
elevations and their proximity to tidal fluctuations.
Vegetation
proximity negatively affected Common Tern hatching success, however vegetation
and elevation co-vary. Since vegetated areas of a shellpile are lower, these
nests were exposed to flooding at a higher frequency than nests on bare shell.
While nest-sites near vegetation may have been selected to provide cover for
eggs and chicks, their proximity to open water caused higher nest failure. At
South Conjers, all but 1 of the 12 nests near vegetation flooded. However,
Gull-billed Terns at Wire Narrows West nested near small plants that had grown
up through the shell, and these nests were significantly more successful than
other nests. For Gull-billed Terns, therefore, the vegetation may have provided
protection and cover, while not exposing the birds to floods.
For
the physical factors involved in hatching success, it appears that elevation
had the strongest effect. As long as nests were low and exposed to flood tides,
the substrate used or the presence of vegetation has little effect on hatching
success. However, if the elevation of a nest is high enough to protect it from
flooding, other factors become more important in explaining hatching success.
Biological and Social Factors
The
effect of clutch size, which may reflect parental age or experience (Nisbet et
al. 1984), on hatching success was not consistent across shellpiles or species.
Clutch size for Common Terns was larger for hatched nests at all shellpiles
except South Conjers, but only significantly so at Wire Narrows West and Man
& Boy. Other factors not analyzed in this paper, such as age of birds or
food availability (Nisbet 1977), may have contributed to this result.
Gull-billed Terns at Wire Narrows West had significantly higher hatching
success with larger clutch sizes. Clutch sizes for all species were similar to
those reported elsewhere (Sears 1978, Erwin 1977b, Nol et al. 1984). At all
shellpiles, Common Terns with 2-egg clutches had the highest hatching success.
This indicates that the investment of energy required to produce two eggs may
be optimal for Common Terns on the shellpiles, or that older, more experienced
birds lay 2-egg clutches.
Predation was, overall, not
a major cause of nest failure at any of the shellpiles except North Conjers
where 64% of early nests were predated. North Conjers had the smallest nesting
colony, with a peak of only 13 nests on June 25 in contrast to Wire Narrows
West that had a peak over 150 nests. Ruddy Turnstones predated over 70% of the
first nests at Wire Narrows East (including 89% of Gull-billed Tern nests) when
the colony had only 17 nests. Burger and Lesser (1979) found that smaller
colonies had higher predation rates, probably because fewer birds were present
to mob predators. However, South Conjers, which was the second smallest colony
but the densest, had the lowest predation rates. Because nests were so dense
over the whole shellpile, mobbing of predators was possibly more effective than
at larger, more spaced colonies. In addition, no other avian species nested on
South Conjers because the island is so small. In contrast, Laughing Gulls,
Herring Gulls, and Greater Black-backed Gulls nested in or around colonies at
Wire Narrows and Man & Boy. However, North and South Conjers islands are
only 300 m apart and so size of colony and density of nests as a factor in warding
off predation may have been the most important factor. As further support for
this, predated Common Tern nests at Wire Narrows West, Wire Narrows East, and
Man & Boy were farther apart than nests not predated (although only
significantly at Wire Narrows West).
Ruddy Turnstones had a
significant impact on hatching success in early-season nests, especially for
Gull-billed Terns at Wire Narrows East. All
observations of Ruddy Turnstones at the shellpiles indicated that Gull-billed
Terns, Common Terns, and Black Skimmers do not treat Ruddy Turnstones as
predators. This observation is similar to others reported earlier (Crossin and
Huber 1971, Parkes et al. 1971, Loftin and Sutton 1979, Brearey and Hilden
1985, Farraway et al. 1986). None of the birds chased the Ruddy Turnstones away
when they were eating eggs.
Hatched
nests were closer together than unsuccessful nests for Common Terns and Black
Skimmers. It is possible that birds nesting in denser areas were able to deter
predators more effectively. However, this relationship only held true at the
Wire Narrows shellpiles. DiCostanzo (1980) found that older birds were the
nucleus of the colony, and it is possible that nests in high density areas were
nests of older, experienced breeders.
Hatching Success Summary
Overall,
hatching success was low on the shellpiles. Erwin (1977b) found hatching
success in Black Skimmers on barrier islands in Virginia to be close to 80%,
while Burger and Gochfeld (1990) found a hatching success rate of 54% on marsh
islands in New Jersey. Only 45% of Black Skimmers nests on shellpiles hatched.
Hatching success for Common Terns in previous studies has ranged from 72% to
88%(Nisbet 1973, Nisbet and Welton 1984). Hatching success for Common Terns was
just over 40% on the shellpiles. Davis et al. (2001) found a hatching success
of 12% for American Oystercatchers in North Carolina. At least 38% of American
Oystercatcher nests hatched on the shellpiles; another 32% had unknown outcomes
and could have hatched. Eyler et al. (1999) found that hatching success for
Gull-billed Terns was lower on marsh islands (54% hatched) than barrier island
beaches (72% hatched). Fifty-four percent of Gull-billed Terns hatched in this
study, although the hatching success was very low (less than 15% hatched) on 2
of the 3 shellpiles they used.
The
multiple regression analysis found, across species and shellpiles, that
elevation and date of nest-initiation explain the most variation in hatching
success. Clutch and nearest-neighbor distance were significant contributors to
the model at some shellpiles, but not all. Since date of nest-initiation
strongly affected hatching success because the floods of early June caused low
nest success in the early season, nest elevation appears to be the most
important and consistent factor explaining hatching success.
Paradoxically,
elevation did not appear to be the most important factor in nest-site
selection, especially for Common Terns. The choice of a nest site by birds
nesting on the shellpiles, therefore, is not based on the factor most important
to reproductive success. The four species involved in this study historically
nested primarily on barrier island beaches until human encroachment, habitat
destruction, and invasion of mammalian predators drove them to seek alternative
nesting sites (Gochfeld 1978, Erwin 1980, Erwin et al. 2001). The selective
pressures determining nest-site selection may be based on selection on barrier
island beaches, and thus may differ from what would be expected on low-lying
marsh islands. The risk of flooding on barrier island beaches is generally
lower relative to marsh islands (Burger and Gochfeld 1991). Eyler et al. (1999)
found lower flooding rates and higher hatching success on barrier island
beaches than on marsh shellpiles. Because flooding due to high tides is
unpredictable and water levels reach different heights each year, the low nest
sites may reflect a trade-off between nesting above flood tide lines and
nesting on preferred substrates.
Sea Level Rise
The
results of this study demonstrate that rising sea levels pose a serious threat
to breeding populations of seabirds on the Eastern Shore of Virginia. If sea
levels rise and no new accumulation of shell on the piles occur, then the
shellpiles will be completely inundated and unsuitable as colony sites. As the
number of floods washing away nests increases over the years, the number of
birds in the colonies will likely decrease. Since barrier island beaches have
become unsuitable colony sites due to the presence of mammalian predators
(Erwin et al. 2001), most breeding habitats in Virginia in the future may be
“sinks” rather than “sources”. However, the shellpiles are storm deposited and
the frequency of storms is predicted to increase with global warming, so new
accumulation of shell may occur.
Management
Implications
The objective of this study
was to determine how experimentally elevating shellpiles affects nest-site
choice and hatching success of the four focal species. I expected higher
elevation sites would be selected for nest-sites because of their increased
protection from flooding, and thus would increase breeding success. The
elevated sites did provide extra protection from flooding; however, the birds
did not select these sites preferentially because factors other than elevation
appeared to be more important in nest-site selection. In order to effectively
manage colony sites to reduce flooding, it is important to understand the
factors important in habitat selection.
Since flooding was the major
cause of nest failure in this study, and because sea levels are expected to
rise, management of low-lying colony sites is still an important objective.
Based on the data collected in this study, a number of improvements to the
design of this project can be made. First, if elevated plots were located in
specific places where birds were known to have nested in previous years, and to
have been flooded, selection of nest-sites on elevated plots may increase. For
example, if an elevated plot was located on the low ridge on Man & Boy where
Gull-billed Terns nested, and flooded, hatching success may improve. An
elevated plot of 20-30 cm would have protected these nests from flooding.
Because plots were located randomly within areas that were used, the plots were
not located where densest aggregation of nests was found. At Wire Narrows West,
an elevated plot in an area where many Common Tern nests failed would only have
to provide 15 cm extra shell height to protect nests from flooding. Larger
elevated plots or plots closer together may also attract more birds because of
the importance of social attraction in nest-site selection for colonial
species. In addition, stabilizing the
elevated plots so that waves cannot erode edges of the plots would help the
success of the plots, especially in the lower-lying areas where many Common
Terns nest.
Second, putting wrack over
selected elevated plots may encourage more birds, especially Common Terns, to
nest at higher elevations. Placing wrack on higher, protected areas of the
shellpile that were not flooded in 2002 may also work to attract birds to
higher sites. At North Conjers wrack covered most of the area used by Common
Terns and this was the only shellpile where the wrack nests had the highest
elevations of all substrates.
Third,
planting of short, sparse vegetation or placement of shade-providing objects on
the shellpile may also attract birds to higher sites. At Wire Narrows West and
East nests surrounded one small plant at a high elevation over the course of
the season (6 nests at WNE, 7 at WNW). At Wire Narrows West the plant died
before the end of the season, but at Wire Narrows East 5 chicks were counted
using the plant as shelter on a hot day. Providing a few small plants or other
sources of shade in key elevated areas may attract birds to more protected
areas.
Finally, if Laughing Gulls deter birds from nesting on more protected sites near the marsh boundary, discouraging nesting of gulls around the shellpile may also increase use of areas of the shellpile not affected by flooding. This may also improve reproductive success if laughing gulls take a significant number of eggs and chicks off the shellpiles.
Conclusions
The
elevation of a nest had the most influence on its success. Despite this, the
majority of nesting pairs of all four species did not select experimentally
elevated plots for nest-sites. Late nests had higher hatching success than
early nests at most shellpiles, primarily because the major flooding occurred
in early June. Predation rates over the course of the season were relatively
even. However, a high percentage of very early nests were destroyed by Ruddy
Turnstone predation. Despite the failure of the elevated sites to attract
nesting birds, the sites were protected from flooding. Because sea level rise
is an imminent threat to nesting waterbirds off the coast of Virginia,
management of nesting sites to reduce flooding is still necessary.
References
Altmann,
J. 1974. Observational study of behavior: Sampling methods. Behaviour
49:227-267.
Breary,
D and O. Hilden. 1985. Nesting and egg-predation by Turnstones Arenaria
interpres in larid colonies. Ornis Scandinavia 16:283-292.
Brown,
R.G.B. Marine birds and climatic warming in the northwest atlantic. 1991.
Canadian Wildlife Service Occasional Paper Number 68:49-54.
Buckley,
F.G. and P. A. Buckley. 1980. Habitat selection and marine birds. Pp 69-112 in
Behavior of marine animals, Volume 4: Marine Birds. (J. Burger, B. Olla, and H.
Winn, eds). Plenum Press, New York, NY.
Buckley,
P.A. and F.G. Buckley. 1984. Seabirds of the North and Middle Atlantic Coast of
the United States: Their status and conservation. Pp 101-133 in Status
and conservation of the world’s seabirds (J. P. Croxall, P. G. H. Evans, and R.
W. Schreiber, eds.). Tech Publ. No. 2., Internl. Council Bird Preserv., Cambridge,
UK.
Burger,
J. 1982. The role of reproductive success in colony-site selection and
abandonment in Black Skimmers (Rynchops niger). Auk 99: 109-115.
Burger,
J. 1985. Habitat selection in marsh-nesting birds. Pages 253-281 In
Habitat selection in birds (Cody, M. ed.). Academic Press, Inc., Orlando, Fl.
Burger,
J. and M. Gochfeld. 1990. The Black Skimmer: Social dynamics of a colonial
species. Columbia University Press, NY.
Burger,
J. and M. Gochfeld. 1991. The Common Tern: Its breeding biology and social
behavior. Columbia University Press, NY.
Burger,
J. and F. Lesser. 1978. Selection of colony sites and nest sites by Common
Terns (Sterna Hirundo) in Ocean County, New Jersey. Ibis 120: 433-449.
Burger,
J. and F. Lesser. 1979. Breeding behavior and success in salt marsh Common Tern
Colonies. Bird-Banding 50: 322-337.
Byrd,
M.A. and D.W. Johnston. 1991. Birds. Pages 477-537 in Virginia’s
endangered species: Proceedings of a symposium (K. Terwilliger, Ed.). McDonald
and Woodward Publishing, Blacksburg, VA.
Clapp,
R.B., D. Morgan-Jacobs, and R.C. Banks. 1983. Marine birds of the southeastern
United States and Gulf of Mexico. Part 3: Charadriiformes. FWS/OBS-83/30, U.S.
Fish and Wildl. Serv., Div. Biol. Serv., Washington, D.C.
Crossin
R.S. and L.N. Huber. 1970. Sooty Tern egg predation by Ruddy Turnstones. Condor
72:372-373.
Davis,
M.A., T.R. Simons, M.J. Groom, J.L. Weaver, and J.R. Cordes. 2001. The Breeding
status of the American Oystercatcher on the east coast of North America and
breeding success in North Carolina. Waterbirds 24: 195-202.
DiCostanzo,
J. 1980. Population dynamics of a common tern colony. Journal of Field
Ornithology 51: 229-243.
Erwin,
R.M. 1977a. Black Skimmer breeding ecology and behavior. Auk 94:709-717.
Erwin,
R.M. 1977b. Foraging and breeding adaptations to different food regimes in
three seabirds: The Common Tern, Sterna hirundo, Royal Tern, Sterna
maxima, and Black Skimmer, Rynchops Niger. Ecology 58: 389-397.
Erwin,
R.M. 1979. Species interactions in a mixed colony of Common Terns (Sterna
hirundo) and Black Skimmers (Rynchops niger). Animal Behavior 27:
1054-1062.
Erwin,
R.M. 1980. Breeding habitat use by colonially nesting waterbirds in two
Mid-Atlantic US Regions under different regimes of human disturbance. Biological
Conservation 18: 39-51.
Erwin,
R.M., and D.C. Smith. 1985. Habitat comparisons and productivity in nesting
common terns on the mid-atlantic coast. Colonial Waterbirds 8:155-165.
Erwin,
R.M., J.D. Nichols, T.B. Eyler, D.B. Stotts, and B.R. Truitt. 1998. Modeling
colony-site dynamics: A case study of Gull-billed Terns (Sterna nilotica)
in coastal Virginia. Auk 155: 970-978.
Erwin
R.M., B.R. Truitt, and J.E. Jimenez. 2001. Ground-nesting waterbirds and
mammalian carnivores in the Virginia barrier island region: Running out of
options. Journal of Coastal Research 17: 292-296.
Erwin,
R.M., D.H. Allen, and D. Jenkins, in press. Created versus natural coastal
islands: Atlantic waterbird populations, habitat choices and management
implications. Estuaries.
ESRI.
2002. ArcGis 8.2. Redlands, California.
Eyler,
T.B., R.M. Erwin, D.B. Stotts, and J.S. Hatfield. 1999. Aspects of hatching
success and chick survival in Gull-billed Terns in coastal Virginia. Waterbirds
22: 54-59.
Farraway, A., J. Thomas, and H. Blokpoel. 1986. Common tern egg predation by ruddy turnstones. Condor 88:521-522.
Fretwell,
S.D. 1972. Populations in a Seasonal Environment. Monographs in Population
Biology, No 5. Princeton University Press, Princeton, N.J.
Gochfeld,
M. 1978. Colony and nest site selection by Black Skimmers. Proceedings of the
Colonial Waterbird Group Conference 1: 78-90.
Gochfeld,
M. and J. Burger. 1994. Black Skimmer (Rynchops niger). In The
birds of North America, No. 108 (A. Poole and F. Gill, eds.). The Academy of
Natural Sciences, Philadelphia and The American Ornithologists’ Union,
Washington, D.C.
Humphrey,
R.C. 1990. Status and range expansion of the American Oystercatcher on the
Atlantic Coast. Transactions Northeast Section Wildlife Society 47: 54-61.
Intergovernmental
Panel on Climate Change, 1996: Climate Change 1995, The science of climate
change. Cambridge University Press, Cambridge, UK.
Jones,
J. 2001. Habitat Selection Studies in Avian Ecology: A Critical Review. Auk
111: 557-562.
Kotliar,
N.B. and J. Burger. 1986. Colony site selection and abandonment by Least Terns
in New Jersey. Biological Conservation 37: 1-21.
Lauro,
B. and J. Burger. 1989. Nest-site selection of American Oystercatchers (Haematopus
palliatus) in salt marshes. Auk 106: 185-192.
Lauro,
B., E. Nol, and M. Vicari. 1992. Nesting density and communal breeding in
American Oystercatchers. The Condor 94:286-289.
Leberg, P.L., P.
Deshotels, S. Pius, and M. Carloss. 1995. Nest sites of seabirds on dredge
islands in coastal Louisiana. Proceedings of the Annual Conference of Southeast
Association of Fish and Wildlife Agencies 49:356-366.
Loftin,
R.W. and S. Sutton.1979. Ruddy Turnstones destroy Royal Tern Colony. Wilson
Bulletin 91:133-135.
Mallach,
T.J. and P.L. Leberg. 1998. Use of dredged material substrates by nesting terns
and Black Skimmers. Journal of Wildlife Management 63:137-146.
Moller,
A.P. 1981. Breeding cycle of the Gull-billed Tern (Gelochelidon nilotica),
especially in relation to colony size. Ardea 69: 193-198.
National
Academy of Sciences 1987. Responding to changes in sea level: Engineering
implications. National Academy Press, Washington D.C.
National
Oceanic and Atmospheric Administration. 2002. Preliminary (tides) water level
data. http://co-ops.nos.noaa.gov/data_res.html.
Nettleship,
D.N., J. Burger, and M. Gochfeld. 1994. Seabirds on islands: Threats, case studies and
action plans: proceedings of the Seabird Specialist Group Workshop held at the
XX World Conference of the International Council for Bird Preservation. Birdlife International,
Cambridge, U.K.
Nisbet,
I.C.T. 1973. Courtship feeding, egg-site, and breeding success in Common Terns.
Nature 241: 141.
Nisbet,
I.C.T. 1977. Courtship Feeding and clutch size in Common Terns. Pages 101-109 in
B. Stonehouse (ed.), Evolutionary Ecology (Biology and Environment vol. 2).
London: Macmillan.
Nisbet,
I.C.T. 1978. Dependence of fledging success on egg size, parental performance,
and egg composition among Common and Roseate Terns, Sterna hirundo and S.
dougallii. Ibis 120:207-214.
Nisbet,
I.C.T. and M.J. Welton. 1984. Seasonal variations in breeding success of Common
Terns: consequences of predation. The Condor 86:53-60.
Nisbet,
I.C.T, J.M. Winchell, and A.E. Hiese. 1984. Influence of age on the breeding
biology of Common Terns. Colonial Waterbirds 7: 117-126.
Nol,
E., A.J. Baker, M.D. Cadman. 1984. Clutch initiation dates, clutch size, and
egg size of the American Oystercatcher in Virginia. The Auk 101: 855-867.
Nol,
E., and R.C. Humphrey. 1994. American Oystercatcher (Haematopus palliatus).
In The birds of North America, No. 82 (A. Poole and F. Gill, eds.). The
Academy of Natural Sciences, Philadelphia and The American Ornithologists’
Union, Washington, D.C.
O’Brien,
R.G. 1998. A tour of UnifyPow: A SAS module/macro for sample-size analysis.
Proceedings of the 23rd SAS Users Group International Conference.
Cary, NC, SAS Institute, 1346-1355.
Parkes
K.C., A, Poole, and H. Lapham 1971. The Ruddy Turnstone as an egg predator.
Wilson Bulletin 83:306-308.
Parnell,
J.F. and R.F. Soots. 1979. Atlas of colonial waterbirds of North Carolina
estuaries. UNC-SG-78-10, North Carolina State University, UNC Sea Grant,
Raleigh, NC.
Parnell,
J.F., R.M. Erwin, and K.C. Molina. 1995. Gull-billed Tern (Sterna nilotica).
In The birds of North America, No. 140 (A. Poole and F. Gill, eds.). The
Academy of Natural Sciences, Philadelphia and The American Ornithologists’
Union, Washington, D.C.
Parnell,
J.F., D.G. Ainley, H. Blockpoel, B. Cain, T.W. Custer, J.L. Dusi, S. Kress,
J.A. Kushlan, W.E. Southern, L.E. Stenzel, and B.C. Thompson.1988. Colonial
waterbird management in North America. Colonial Waterbirds 11: 129-168.
Pius,
S.M. and P.L. Leberg. 1997. Aggression and nest spacing in single and mixed
species groups of seabirds. Oecologia 111: 144-150.
SAS
Institute, Inc. 1991. SAS/STAT user’s guide release 8.2. SAS Institute, Inc.,
Cary, NC.
Sears,
H.F. 1978. Nesting behavior of the Gull-billed Tern. Bird Banding 49: 1-15.
Shealer,
D.A. and S.W. Kress. 1991. Nocturnal abandonment response to Black-crowned
Night Heron disturbance in a Common Tern colony. Colonial Waterbirds 14:51-56.
Spendelow,
J.A. 1982. An analysis of temporal variation in, and the effects of habitat
modification on, the reproductive success of Roseate Terns. Colonial Waterbirds
5:19-31.
SPSS,
Inc. 2001. SPSS 11.0. Chicago, Il.
Via,
J. and D.C. Duffy. 1992. Gull-billed Tern (Sterna nilotica). Pages
135-148 in Migratory Nongame Birds of Management Concern in the
Northeast. U.S. Dep. Inter., Fish and Wildl. Serv., Newton Corner,
Massachusetts. 400pp.
Virginia
Institute of Marine Science, 2002. Tide calendars for 2002, Wachapreague.
http://www.vims.edu/resources/tides/2002.html. Gloucester, VA.
Williams, B., R. Beck, B. Akers, and J. Via. 1990.
Longitudinal surveys of the beach nesting and colonial waterbirds of the
Virginia Barrier Islands. Virginia Journal of Science 41: 381-388.
Appendix 3. Differences in nest
elevations between species at each shellpile. a: WNW ANOVA F=54.65, P<0.001.
GBTE higher than BLSK and COTE, AMOY higher than COTE, and BLSK higher
than COTE all at P=0.05 using Tukey’s studentized range test. b: WNE ANOVA, F=11.38, P<0.001. BLSK and GBTE
higher than COTE at P=0.05 using Tukey’s studentized range test. c: MB T=2.08, P=0.04, GBTE lower than
COTE. d: NC T=-3.15, P=0.002, COTE higher
than AMOY.