VCR_LTER_III
The central
hypothesis of the VCR LTER is that ecosystem, landscape and successional
patterns are controlled by the relative vertical positions of the land,
sea, and fresh-water table. Large transient and small progressive changes
in the position of these free surfaces result in disturbance of ecosystem
processes and in physiological stresses that may lead to system state change.
Variations in the elevations of these critical surfaces result from local
weather or climate change, such as short-term storm-generated fluctuation
in sea/land levels or long-term sea level rise. Ecological processes, including
species extinctions and invasions that alter rates of erosion and deposition,
also affect the positions of these free surfaces relative to one another.
In many instances, the joint effects of contemporaneous disturbances at
different temporal and spatial scales result in state changes.
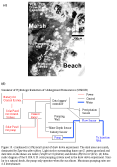
The Virginia Coast Reserve (VCR) is a
complex assemblage of 14 barrier islands, associated inlets and beaches,
extensive back barrier islands, shallow bays and deep channels, mud flats,
expansive salt marshes, and contiguous mainland fringing marshes, deciduous
and evergreen forests, and agricultural fields (see Fig. 1
and 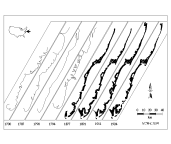 Fig. 6 ).
It extends 110 km along the seaward margin of the southern Delmarva Peninsula.
The low-lying landscape (mean elevation < 2 m) provides an unusually
sensitive location for studying and monitoring ecosystem dynamics and the
effects of both atmospheric and oceanic climate variability on a range
of natural ecosystems. The 14,000 ha site is part of the mid-Atlantic coastal
plain which extends from the fall-line 100 km inland to the edge of the
continental shelf. The plain has a seaward slope of only 0.1%.Where sea
level intersects this sloping plain, small changes in elevation of either
the land or the sea result in large horizontal changes that determine landscape
patterns and ecosystem structure and dynamics.
Fig. 6 ).
It extends 110 km along the seaward margin of the southern Delmarva Peninsula.
The low-lying landscape (mean elevation < 2 m) provides an unusually
sensitive location for studying and monitoring ecosystem dynamics and the
effects of both atmospheric and oceanic climate variability on a range
of natural ecosystems. The 14,000 ha site is part of the mid-Atlantic coastal
plain which extends from the fall-line 100 km inland to the edge of the
continental shelf. The plain has a seaward slope of only 0.1%.Where sea
level intersects this sloping plain, small changes in elevation of either
the land or the sea result in large horizontal changes that determine landscape
patterns and ecosystem structure and dynamics.
The vegetation of the
islands, marshes, and the mainland-lagoon interface is conspicuously zonal
(McCaffrey and Dueser 1990a, b, Ehrenfeld 1990) in character with sub-scale
patchiness and sharp transitions between patches. High and low salt marshes,
unvegetated mud flats, grasslands, shrub savannas and maritime forests
occur in close proximity, with sharp ecotones between adjacent ecosystems.
Vegetation heterogeneity is further accentuated by interannual climate
variation: precipitation records indicate a range of between 85 and 140
cm yr-1 (Hayden 1979; Bolyard et al. 1979). This variation
arises primarily from variation in summer thunderstorm rainfall and depends,
in part, on local land and sea temperature differences. Further, 45% of
late summer and autumn rainfall comes from tropical storms (Hayden 1979).
The VCR is an extremely dynamic, regularly disturbed landscape comprising
elements (forests, marshes, sand dunes, grasslands, etc.) that differ in
degrees normally associated with biome-level differences. The types of
ecosystem changes that occur spatially across continental biomes and temporally
over glacial and interglacial episodes happen on decadal time scales at
the VCR. Successional chronosequences in accretionary island landscapes
are well documented, and succession proceeds rapidly (Hayden et al. 1991).
Ecosystem dynamics at the local level produce landscape structure at higher
levels of organization. Yet for all of this disturbance and change, or
perhaps because of it, the landscape is comprised of repeating units and
is seemingly self-ordered. The barrier islands form a gentle arc with the
regularity of beads on a string. The morphologies of barrier islands, and
their lagoons and mainland interface, repeat in form and dimension. Ecosystem
state variables exhibit significant decadal and century-long trends that
are driven by extreme perturbations at meteorological and climatological
time scales.
The climate of the VCR is dominated by coastal storms
or Northeasters (Dolan et al. 1987) 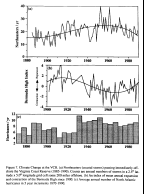 (Fig. 7a)
and the Bermuda High pressure system (Davis et al. 1994) (Fig. 7b). Each year an average of 38 extratropical
storms occur with magnitudes sufficient to rework beach sands and to elevate
tides above astronomical norms (Hayden 1976 1981; Dolan et al. 1987 1988).
These storms generate waves from the northeast and drive sand southward
along the coast. The Bermuda High pressure system generates southeast swell
and drives sediment northward along the coast. Like the Northeasters and
the size of the Bermuda High, hurricanes have also exhibited long term
trends (Fig. 7c). These storms are largely responsible for the production
of waves and surges which in turn change the morphology of the islands
(Dolan et al. 1979), the associated vegetative cover, and ultimately in
combination with sea level rise, the landward migration of islands across
lagoonal marshes (Hayden et al. 1980, 1981) and the encroachment of marshes
into the forests on the mainland (See Fig. 3B, page RPS 3) (Brinson et
al. 1994). In effect, the site is a perpetual experiment in ecosystem disturbance,
plant succession and landscape structuring. Harris (1992) determined that
90% of the landscape of Hog Island (our principal study site for barrier
islands) seaward of its fringing marsh dates from 1871 or later (see Fig. 2a,
page RPS 2). The extent of marshes throughout the VCR declined 11%
from 1852 to 1960 (Knowlton 1971). Farm fences on hypersaline marshes are
evidence of the encroachment of saline water into fields and fresh-water
marshes along the lagoon mainland interface.
(Fig. 7a)
and the Bermuda High pressure system (Davis et al. 1994) (Fig. 7b). Each year an average of 38 extratropical
storms occur with magnitudes sufficient to rework beach sands and to elevate
tides above astronomical norms (Hayden 1976 1981; Dolan et al. 1987 1988).
These storms generate waves from the northeast and drive sand southward
along the coast. The Bermuda High pressure system generates southeast swell
and drives sediment northward along the coast. Like the Northeasters and
the size of the Bermuda High, hurricanes have also exhibited long term
trends (Fig. 7c). These storms are largely responsible for the production
of waves and surges which in turn change the morphology of the islands
(Dolan et al. 1979), the associated vegetative cover, and ultimately in
combination with sea level rise, the landward migration of islands across
lagoonal marshes (Hayden et al. 1980, 1981) and the encroachment of marshes
into the forests on the mainland (See Fig. 3B, page RPS 3) (Brinson et
al. 1994). In effect, the site is a perpetual experiment in ecosystem disturbance,
plant succession and landscape structuring. Harris (1992) determined that
90% of the landscape of Hog Island (our principal study site for barrier
islands) seaward of its fringing marsh dates from 1871 or later (see Fig. 2a,
page RPS 2). The extent of marshes throughout the VCR declined 11%
from 1852 to 1960 (Knowlton 1971). Farm fences on hypersaline marshes are
evidence of the encroachment of saline water into fields and fresh-water
marshes along the lagoon mainland interface.
The "Great Halloween
Storm of 1991" and its effect on the VCR landscape illustrates the
exceptionally dynamic nature of this landscape. On October 30, 1991, the
VCR was hit by a storm that evolved from an extratropical storm (a "northeaster")
moving eastward to merge with a passing tropical storm, Grace. The resultant
Great Halloween Storm (Dolan and Davis 1992) was a major coastal-storm
event from Cape Hatteras, N. C. to Kennebunkport, Me. The storm remained
offshore and produced considerable damage even at a distance. At the VCR,
waves at sea reached heights over 11 m and the eastern fronts of the barrier
islands were pounded with waves of 3 - 5 m. This was the most powerful
storm of the 1447 coastal storms in our LTER data archives (1942-1994).
Statistical estimations of its probability indicate a 1000 year plus return
interval, i.e., this storm was exceptional. The Great Halloween Storm is
comparable to historical disturbance events -- The Great Storm of 1667,
The Accomac Hurricane of 1693, the hurricanes of 1749 and 1933. Each produced
similar waves and still water levels 3 to 5 m above normal. Immediately
following the Halloween Storm, ground and aircraft reconnaissance of the
site indicated that in some locations 10s of meters of beach were removed
from Hog Island. The inlet between Parramore and Crescent Islands was filled
with sand. Marshes behind Parramore Island were buried by as much as 1
m of sand. Large sections of Parramore Island's south end eroded away.
Fresh water wells in transects across Hog island became saline. The Halloween
Storm and other large storms leave extensive deposits of wrack that appear
to be partly responsible for patchiness of marsh vegetation (Brinson et
al. 1994). Several new marsh surfaces developed on which we have installed
an extensive array of permanent plots and transects to chart the colonization
and development of a salt marshes on newly deposited substrates.
Changes in the states of VCR ecosystems
can be dramatic. Tens of thousands of years ago, our site was a maritime
boreal forest of Pinus strobus and Picea sp. (Hayden 1990,
Bonan and Hayden 1990a, b; Emory et al. 1967). Seven thousand years ago,
what are now open lagoons were forested uplands with stream channel estuaries.
Five thousand years ago, these sites changed to tidal channels and marshes
(Newman and Munsart 1968). In the last 1500 years the site changed to today's
lagoons, marshes and barrier islands perched on the much older surface
(Oertel et al. 1989a, c, and Harrison et al. 1965).
The contemporary
barrier island-lagoon-mainland complex took form during the late Holocene
rise in sea level, although the underlying topographic framework can be
traced back to earlier high stands of sea level during the Pleistocene
(Oertel et al. 1989a). Rapid change has taken place during the last few
thousand years with the barrier island complex migrating westward across
the continental slope at a rate as high as 10-3 km yr-1
(Finkelstein et al. 1987). The modern islands typically exhibit shoreline
change characterized by lateral accretion and erosion as high as 13 m yr-1 (Dolan et al. 1979) creating one of the most dynamic coasts
in the United States.
While the entire site has undergone a sequence
of state changes over the Holocene, landscape/ecosystem elements within
the site have undergone equally large state changes within the present
century. Mockhorn Island, once agricultural land and formerly forest, has
been abandoned and is now a very young salt marsh. At the turn of the century,
the salt marshes that now fringe the mainland were fenced, upland pastures.The
bay bottoms, which were eel grass meadows at the turn of the century, are
now unvegetated bottoms (Hayden et al. 1991). At the time of settlement
of Hog Island's town of Broadwater in the mid 1800s, the south end of Hog
Island was forested. It is now an overwash-flat grassland (Rice et al.
1976, Fitch 1991, Hayden et al. 1980). The north end of Hog Island was
several hundred meters wide and consisted of a narrow beach affronting
a marsh in 1852; and, it is now a complex chronosequence of ridges and
swales nearly 1.5 km wide (Hayden et al. 1991). The VCR is a mosaic of
landscapes and ecotones with rates of change exceeded in North America
only by the coastal margin of the Mississippi Delta (Dolan et al. 1982).
H.C. Cowles (1899) once characterized ecological succession as "a
variable approaching a variable" indicating that internal ecosystem
processes (the first "variable") interacted to produce structural
dynamics that seem directed toward a configuration that changes with the
environment (a moving target and the second "variable"). At the
VCR, transitions between ecosystem states can occur with exceptional abruptness.
We envision the ecosystem dynamics at the VCR as having a tendency to be
self-ordering or successional within limits. Under certain intensities
of disturbance, systems are displaced sufficiently that their dynamics
track a different "target" or state (Hayden et al. 1991) 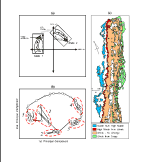 (Fig. 8a & 8b). For example, islands with maritime forests, like the south
end of Hog Island in the 1920s and Parramore Island today, experience forest
die back following shoreline erosion, loss of island elevation, and depletion
freshwater reserves necessary for forest maintenance. Unless new island
building takes place, a replacement forest will not result. Rising sea
level is currently forcing state changes on the barrier islands including
high marsh replacing shrubs and low marsh replacing high marsh (Fig. 8c).
(Fig. 8a & 8b). For example, islands with maritime forests, like the south
end of Hog Island in the 1920s and Parramore Island today, experience forest
die back following shoreline erosion, loss of island elevation, and depletion
freshwater reserves necessary for forest maintenance. Unless new island
building takes place, a replacement forest will not result. Rising sea
level is currently forcing state changes on the barrier islands including
high marsh replacing shrubs and low marsh replacing high marsh (Fig. 8c).
There are two elements to our theoretical conception of landscape dynamics
on the VCR LTER. First, there are ecosystem states, in a multivariate Hutchinsonian
sense, within which successional dynamics operate. Second, there are events
that produce transitions from one of these states to another (Fig. 8a &
8b). At the VCR, one can see both successional dynamics within these ecosystem
states and transitional dynamics between given ecosystem states. In spite
of this early stage in theoretical development of this notion, we are convinced
the observational and multivariate definition of ecosystem states is possible
in the sense of Allen et al. (1977), Birks and Berglund (1979), Lamb (1985),
and Jacobson and Grimm (1986) (Fig. 8b). Our concept of landscape dynamics
(Hayden et al. 1991) is based on observations of the dynamics in reconstructions
of landscape change over 1000s of years based on pollen records (Emory
et al. 1967), from landscape reconstruction from 40 years of aerial photos
(Dolan et al. 1979), and from historical accounts and survey maps (Rice
et al. 1976). We can infer such dynamics from field evidence, for example,
dead tree trunks in marshes and marsh peats on beaches. We feel this class
of dynamics, while frequent and dramatic, is not unique to our site and
is relatively widespread in a range of ecosystems. These dynamics occur
over time intervals that are not usually subject to direct observation
or experimentation, at the VCR these dynamics are rapid and accessible
through long-term research.
We view the dominant cause of ecosystem
state change at the VCR, and also the initiation of new successional sequences
within states, as arising from vertical changes in at least one of three
free surfaces: the land, sea, and fresh-water table. We use the term free
surface rather than simply surface to emphasize that these are not
static surfaces. Mass and energy are exchanged across these surfaces and
the elevations of these surfaces exhibit long-term trends, periodic oscillations
and episodic disturbances. Variations in the elevations of land, sea and
water table surfaces arise from relative sea-level rise, storm-generated
changes in levels of the sea and the land, and ecological processes including
species extinctions. Our long-term experiments and long-term observation
and monitoring programs focus on these free surfaces and on the role of
they play in ecosystem succession and state changes.
The concept of free surfaces, as we use it,
is the same as used in hydrology and meteorology. Our hypotheses focus
on the relationships of three free surfaces: fresh-water table, land and
sea level 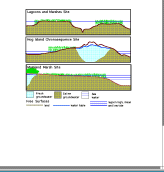 (Fig. 9). While other free surfaces exist, e.g., the free surface
characterized by the sediment oxic/anoxic interface, we focus on these
three because studies over the last 6 years suggest that the VCR LTER site
is structured to a large extent by these surfaces. At the VCR small variations
in elevations or slopes of the fresh-water table, land or sea level surfaces
can result in ecosystem and landscape changes equivalent to continental
biome transitions . Aeolian transport of sand inland from the beach may
result in higher base island elevations and in the formation of dunes.
It is the base elevation of the land above the extremes of the astronomical
tides that permits significant trapping of rainwater and the development
of a perched freshwater lens which is the confined fresh water resource
for terrestrial vegetation (Bolyard et al. 1979). The storage capacity
for this resource depends upon the difference between the elevation of
the fresh-water table and on the elevation of the land free surface (Ghyben-Herzberg
principle) and on local precipitation and evaporation. Where the fresh
water storage capacity is large, forests with high leaf area index (LAI)
may develop; where the resource is modest, lower LAI grasslands are common;
and, where the land level approaches the level of high tides and where
fresh water reserves are not present, infrequent tidal flooding combined
with high evaporation rates may result in hypersaline salt flats or Salicornia
virginica-dominated zones.
(Fig. 9). While other free surfaces exist, e.g., the free surface
characterized by the sediment oxic/anoxic interface, we focus on these
three because studies over the last 6 years suggest that the VCR LTER site
is structured to a large extent by these surfaces. At the VCR small variations
in elevations or slopes of the fresh-water table, land or sea level surfaces
can result in ecosystem and landscape changes equivalent to continental
biome transitions . Aeolian transport of sand inland from the beach may
result in higher base island elevations and in the formation of dunes.
It is the base elevation of the land above the extremes of the astronomical
tides that permits significant trapping of rainwater and the development
of a perched freshwater lens which is the confined fresh water resource
for terrestrial vegetation (Bolyard et al. 1979). The storage capacity
for this resource depends upon the difference between the elevation of
the fresh-water table and on the elevation of the land free surface (Ghyben-Herzberg
principle) and on local precipitation and evaporation. Where the fresh
water storage capacity is large, forests with high leaf area index (LAI)
may develop; where the resource is modest, lower LAI grasslands are common;
and, where the land level approaches the level of high tides and where
fresh water reserves are not present, infrequent tidal flooding combined
with high evaporation rates may result in hypersaline salt flats or Salicornia
virginica-dominated zones.
Neither the level of the land nor the
level of the sea is stationary (Table 2). At the south end of the VCR,
the land is subsiding around 2 mm yr-1 and the eustatic level
of the sea is rising at about 1 mm yr-1 giving a relative
sea level rise of 3.0 mm yr-1 (Emory and Aubrey 1991). Oertel
(1989b) places the relative rate of sea level rise (eustatic rise + land
subsidence) between 2.8 to 4.2 mm yr-1. The rate of relative
sea level rise decreases toward the mainland as well as southward and northward
for hundreds of kilometers (Emory and Aubrey 1991). In general, this rate
of relative sea level rise exceeds the capacity of marshes to build upward
unless marine sands or terrestrial sediments are readily available for
deposition (Ray and Hayden 1992; Kastler 1993) or unless biogenic accretion
can keep pace with the rising sea (Brinson et al. 1994). Land level and
eustatic sea-level surfaces, measured locally as the single parameter relative
sea level, are important free surfaces for VCR ecosystems. When changes
in land and sea level elevations exceed a threshold, system state change
occur (Ray et al. 1992). The characteristic temporal variability of these
critical state variables is summarized in Table 2.
Locally, the elevation
of the land free surface is controlled by sedimentation and erosion of
marine and terrestrial mineral sediments and organic matter accumulation
(Hackney and Cleary 1987; Brinson et al. 1994). For VCR ecosystems, increases
in elevation due to sedimentation may offset the regional subsidence of
the land and eustatic rise of the sea. Additionally, because most of the
inorganic sedimentation above mean sea level is transported hydraulically,
water levels during northeasters and tropical storms are the critical agents
of change raising the land surface elevations above the levels of the astronomical
tides (Dolan and Godfrey 1973). Below ground organic matter accumulation
(Blum 1993) and overwash of marine sands (Kochel and Wampfler 1989) are
also critical processes at the VCR that alter land elevations and may give
rise to system state changes.
These considerations have led us to begin
long-term experiments which manipulate critical free surfaces. The first
two involve direct manipulation of the pattern of inundation frequency
and magnitude for two types of marsh (fringe and insular-lagoon), using
approaches appropriate for the marsh type, to provide insight into the
response of marshes to sea level rise. The third experiment employs a long-term
manipulation of the unconfined freshwater lens on a coastal barrier island
to study state changes arising from climate change and to study ecosystem-level
water relations. These experiments are designed to build on and to focus
our earlier research findings, and to emphasize the free surface controlling
factors of water table, land, and sea elevations and to understand the
biogeographic organization of the VCR.
The Hypotheses. The research programs of the VCR LTER
are focused on understanding the relationships between slow, long-term
changes in critical free surfaces and rapid, short-term disturbances in
these same surfaces that trigger ecosystem state changes and set in motion
or alter successional sequences 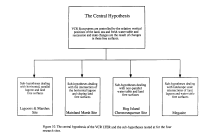 (Fig. 10).
(Fig. 10).
The central
hypothesis of the VCR LTER is that ecosystem, landscape and successional
patterns are controlled by the relative vertical positions of the land,
sea, and fresh-water table. Large transient and small progressive changes
in the position of these free surfaces result in disturbance of ecosystem
processes and in physiological stresses that may lead to system state change.
Variations in the elevations of these critical surfaces result from local
weather or climate change, such as short-term storm-generated fluctuation
in sea/land levels or long-term sea level rise. Ecological processes, including
species extinctions and invasions that alter rates of erosion and deposition,
also affect the positions of these free surfaces relative to one another.
In many instances, the joint effects of contemporaneous disturbances at
different temporal and spatial scales result in state changes.
Several types of relationships between the three free surfaces occur at
the VCR (Figure 10 and 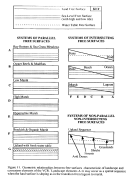 Fig. 11). An understanding of these relationships
goes directly to the theoretical basis for the sub-hypotheses that guide
our field studies. Nine relationships are detailed below. At any one location
within the VCR a subset of these relationships apply. We do not refer to
these relationships as hypotheses as, in their general formulation, they
are not the object of our field studies. Nonetheless, the sub-hypotheses
of our central hypothesis are founded from these relationships.
Fig. 11). An understanding of these relationships
goes directly to the theoretical basis for the sub-hypotheses that guide
our field studies. Nine relationships are detailed below. At any one location
within the VCR a subset of these relationships apply. We do not refer to
these relationships as hypotheses as, in their general formulation, they
are not the object of our field studies. Nonetheless, the sub-hypotheses
of our central hypothesis are founded from these relationships.
- 1. Long-term (> 1 yr) changes in the mean elevation of a horizontal
free surface intersecting a sloping free surface causes associated ecotones
to shift position horizontally.
- 2. Long-term (> 1 yr) changes
in the mean elevation of a free surface parallel to a static free surface
causes an ecosystem state change.
- 3. Short-term (<= 1 yr) periodicities
in the elevation of a free surface relative to parallel, static free surface
gives rise to a characteristic ecosystem type which depends on the frequency
of surface elevation variation, the amplitude of the free surface elevation
change, and the relative position of the static free surface.
- 4.
A horizontal gradient in the mean elevation of one free surface relative
to a horizontal second free surface results in horizontal zonation of ecosystems.
A change in magnitude of the gradient in elevation compresses or expands
the zonal arrangements of ecosystems. A change in the elevation of the
horizontal surface displaces the zonal system of ecosystems horizontally.
- 5. Long-term maintenance of ecosystem structure, in an environment
with free surface elevation changes, requires parallel changes in at least
one of the other free surfaces. If these conditions are not met, an ecosystem
state arises, a results that could not be achieved by autogenic succession.
- 6. Patchiness in community structure arises from a spatially
commensurate variation in the elevation of one or more free surfaces. The
land free surface elevation heterogeneity is a primary source of patchiness
at a number of spatial scales at the VCR.
- 7. Disturbances that
change the geometry of free surfaces may cause ecosystem state change.
A disturbance that does not change the geometry of the free surfaces does
not cause an ecosystem state change, but may set the system back to an
earlier successional stage.
- 8. Nutrient cycling, above and below
ground production, decomposition, and local trophic structure are directly
or indirectly dependent on positional inter-relationships of the free surfaces
associated with the specific ecosystem.
- 9. Keystone species at
the VCR are those that have the capacity to alter the elevations of the
three free surfaces. Ammophila on islands, Spartina on marshes,
oysters on mudflats, and sea grass on bay bottoms all influence the deposition
or erosion of sediments.
Our research program is designed to
investigate relationships between the three free surfaces and the ecosystems
that depend on them. We have also begun three long-term manipulative experiments
that directly alter critical free surfaces. We will monitor the long-term
ecosystem changes at each experimental site. Additionally, we have paired
each experiment with landscape units which have, over the last several
decades, experienced natural variations in the same free surfaces. Our
approach during the past 6.5 years has been to study the controls on succession
and state changes at four research venues within the Virginia Coast Reserve:
the VCR Megasite, the North Hog Chronosequence Site, the Hog Island Bay
Lagoon and Mainland Marsh Site. At each site we are engaged in core-area
work that tests our research hypotheses, specifically supports our experimental
efforts, provides critical data for models, and provides useful information
for the management of the site by our host, The Nature Conservancy. At
our research locations, we are investigating the nine aspects of our central
hypothesis. At the VCR Megasite we focus our research on the intersections
of the three free surfaces at large geographic and long time scales. At
the North Hog Chronosequence Site we focus on the non-parallel fresh-water
table free surface and its elevation variations relative to the land free
surface across a 120 year chronosequence of ecosystems. The marsh-land
surface and its interactions with the parallel yet oscillatory lagoon water
free surface is the focus of research at the Hog Island Bay Lagoons and
Marshes Site. At the Mainland-Marsh Site we focus on the oscillatory free
surface of the level of the lagoon intersecting the sloping land and water
table free surfaces. We rely on imagery, networks of sensors and permanent
plots, wide-area surveys, transects, and reconnaissance programs as well
as sediment coring programs to deal with historical and paleoecological
variations in VCR landscapes.
The Megasite consists of The Nature Conservancy\qs Virginia Coast Reserve
and the lands westward to the watershed divide of the Delmarva Peninsula
(see Figure 1). The major boundaries of this strikingly zonal and patchy
landscape arise from the separation between free surfaces and intersections
of two or three critical free surfaces and gives rise to rapidly changing
ecosystems 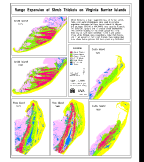 (Fig. 12). The zones and patches which comprise the VCR have
characteristic relationships among the three free surfaces. The central
hypothesis as applied to the Megasite is as follows:
(Fig. 12). The zones and patches which comprise the VCR have
characteristic relationships among the three free surfaces. The central
hypothesis as applied to the Megasite is as follows:
Long-term
progressive and short-term disturbance-driven changes in the intersections
of land, lagoon and water-table free surfaces of the VCR cause changes
in the spatial (10s m to 10s km) organization of VCR ecosystems and shifts
in ecotones.
Sub-hypotheses being studied at the Megasite (MS)
include:
- The sedimentary layer produced by the 1933 hurricane
that buried then-existing marshes is present throughout Hog Island Bay
and is thickest where sea grass meadows and were extensive before 1933.(Zieman
and Hayden)
- The shorezone of the barrier islands switches from
accretional to erosional with a return period of approximately 80 years.
We further hypothesize that the transition from accretion to erosion patterns
is an Atlantic Coast-wide phenomenon that is caused by changes in winter
storm(s) climate and long-term changes in the subtropical anticyclone of
the North Atlantic. (Hayden and Dolan).
- Seaside vegetation composition
on the barrier islands is a response to winter storm overwash disturbances
(changes in land free surface elevation) and marine flooding; the effects
vary depending on whether sections of the island are erosional or accretional.
(Hayden and Shugart).
- Elevation of the land free surface above
sea and fresh water surfaces are major determinants of the capacity of
an island to support viable populations of vertebrates. Extinctions of
vertebrate populations may result from changes in one or more of these
surfaces. (Dueser, Porter and Moncrief).
- Reworking of regressive
strandplain land free surfaces during the late Holocene transgression produced
a characteristic stratigraphy due to the release of "fines" into
the water column which increased lagoonal turbidity. The "fines"
are preferentially deposited in wave-protected areas, and the marshes of
small watersheds on the lagoon margins. (Oertel)
- Lagoonal waters
are presently light-limited and turbid as the result of both inorganic
and organic (high C/N values) particulate suspension. Increased primary
production by phytoplankton and an increase in macroalgae will reduce organic
turbidity in lagoonal waters. It is further hypothesized that this increase
in clarity will be a key factor in the return of seagrasses to the VCR.
( Zieman and Hayden).
Megasite studies depend upon repeated
synoptic surveys and measurements across a wide geographic area and at
time intervals appropriate to determining the long-term dynamics. The benchmark
Megasite inventory is Dueser et al (1976). Time intervals for such surveys
range from 1 to 5 years, depending on our understanding of the dynamics
of the system under study and the occurrence of significant disturbances.
Spatial density of sampling depends on the type of data and the structure
of the landscape. An inventory of long-term records is provided in Section
2: Long-term Data Sets. We are initiating an extensive survey of marsh
surface coring to measure the thickness and position in the stratigraphic
column of sedimentary deposits that we hypothesize were laid down in 1933
(MS Ho-1). We have dated carbon from this layer using high-sensitivity
accelerator mass spectrometry radio-carbon dating and place it in the 1930s.
The survey of this stratigraphic surface will provide a widespread time
marker for many studies.
Megasite programs focus on recording elevations
of critical free surfaces and ecosystem attributes. Studies of land surface
elevations focus on measurements of changes in the shorezone. Within this
zone, island building and destruction, and the reworking of the island
sediments are driven by storm waves and surges. Surveys of the shorezone,
the intersection of the sea-level and land free surfaces, rely on aerial
photography (1949-1993). Beginning in 1994 we will use GPS surveys to determine
shoreline and ecotone positions. This will permit improved schedules for
surveys and reduce post-processing requirements. Storm and wave climate
data from the National Weather Service is added to our data archives monthly(1885-1994).
Studies to date, have shown that the shorezone of Hog Island has undergon
fundamental reversal in the direction of change in 1967 (Fenster et al.
1994)(MS Ho-2). Reversals have occurred once or twice per century for the
last several hundred years (Harris 1992). Our current interpretation is
that winter storms and summer swell from the Bermuda high pressure cell
(see Fig. 7, page PD 2) have changed systematically over the last century
(Hayden 1981, 1982; Davis et al. 1994). The specific mechanism involved
is hypothesized to be a change in the net along-the-coast direction of
movement of sediments (MS Ho-2). Along the island where the shorezone is
widest, seaward terrestrial environments are flooded with saline water
during coastal storms, and may be buried by sands transported from the
beach. We have mapped the probabilities of such burials on Hog Island and
have determined for each probability class the plant species which survived
in that disturbance zone (Fahrig, Hayden and Dolan 1993)(see Fig. 5, page
RPS 5). During the renewal period we will verify the results of this work
(MS Ho-3) on other islands and will use this survival vs. disturbance probability
information in our model ISLAND (see Section 2: Models and Synthesis page
PD 23).
Work on MS Ho-4 examines the causes of extinction and immigration
by linking multiannual surveys of multiple islands (varying in elevation),
semi-annual censuses of small mammal populations on four transects on Hog
Island (varying in availability of fresh water and probability of overwash),
intensive observations of inter-island movements (Forys and Dueser 1993)
and genetic (mitochondrial DNA, allozyme and microsatellite DNA) analyses
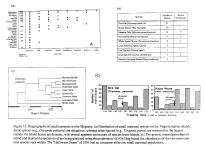 (Fig. 13). Extensive multi-island surveys were completed in 1988-1989 and
1993. We are examining relationships between insular populations using
genetic analyses of samples from 11 islands sampled in 1993. Aerial photo
and map surveys indicate an 11% loss of lagoon and back-barrier marshes
since 1852 (Knowlton 1971). More recent surveys covering the last 30 years
(Kastler 1993), found that loss of island marsh exceeds development of
new marsh while mainland marsh has increased uby 8%. We hypothesize (MS
Ho-5) that the lagoons are enriched in low-nitrogen carbon particles winnowed
from of the eroded marshes. We will quantify loading of lagoon waters with
low quality carbon and new sediments from marshes. In conjunction with
the US MAB program, we have initiated, a study of the role of nutrient
enriched groundwater from the mainland which may be the source of the high
levels of nutrients in the creek and lagoon waters. General observations
indicate that lagoon bottoms that were seagrass meadows prior to the extinction
of the eel grass in the early 1930s are becoming especially rich in macro-algae
(MS Ho-6). Transects, used in preliminary work, together with new central
lagoon permanent plots will be established and used to measure changes
in macroalgal biomass and community composition.
(Fig. 13). Extensive multi-island surveys were completed in 1988-1989 and
1993. We are examining relationships between insular populations using
genetic analyses of samples from 11 islands sampled in 1993. Aerial photo
and map surveys indicate an 11% loss of lagoon and back-barrier marshes
since 1852 (Knowlton 1971). More recent surveys covering the last 30 years
(Kastler 1993), found that loss of island marsh exceeds development of
new marsh while mainland marsh has increased uby 8%. We hypothesize (MS
Ho-5) that the lagoons are enriched in low-nitrogen carbon particles winnowed
from of the eroded marshes. We will quantify loading of lagoon waters with
low quality carbon and new sediments from marshes. In conjunction with
the US MAB program, we have initiated, a study of the role of nutrient
enriched groundwater from the mainland which may be the source of the high
levels of nutrients in the creek and lagoon waters. General observations
indicate that lagoon bottoms that were seagrass meadows prior to the extinction
of the eel grass in the early 1930s are becoming especially rich in macro-algae
(MS Ho-6). Transects, used in preliminary work, together with new central
lagoon permanent plots will be established and used to measure changes
in macroalgal biomass and community composition.
Research infrastructure
of the VCR Megasite includes: 1) a 60-monument, cm-level GPS network, 2)
a system of dated landscapes, 3) a network of surveyed overwash terrace,
4) a network of sediment cores, 5) a benchmark system of vegetation plots,
6) a system of mammal transects on 23 island and marsh surfaces, 7) remote
sensing calibration facilities, 8) a mm-level set of benchmarks at research
sites, and 9) a geo-rectified, 50-meter interval VCR-wide base map system
for shorezone landscape studies. These on-site resources for site-wide
research are supported by a remote sensing laboratory in Charlottesville.
The water-table free surface and the land free surface are neither parallel
nor horizontal on the barrier islands. The spatial and temporal variations
in relationships of these two free surfaces control ecosystem structure,
dynamics and long-term changes. The central hypothesis applied to the North
Hog Island Chronosequence site  (Fig. 14) is:
(Fig. 14) is:
Terrestrial vegetation
on coastal barrier islands is limited primarily by the relationship of
the non-parallel land and water table free surfaces. The relative elevation
of the fresh-water table and the magnitude of the reserve of fresh water
available structures the vegetation of the barrier islands.
Working
sub-hypotheses for North Hog Island (HI) Chronosequence include:
- The depth of the fresh water table below the surface of the
island depends on the elevation of the land above mean sea level. (Kochel,
Hayden, Furman, and Porter)
- In stable or accreting regions of
the islands, species composition of the vegetation is determined by island
elevation, depth to the freshwater table, and variations in water table
elevation due to rainwater input and evapotranspiration losses. (Brinson,
Porter and Hayden)
- In unstable regions of the island, plant species
composition and successional processes are determined by depth to the freshwater
table, climate variations and by the frequency and magnitude of disturbance
from coastal storms. (Young and Hayden)
- Accreting regions of the
islands have landscapes ordered in age from the sea landward. Along this
chronosequence the relative positions of the land and water-table free
surfaces have been in place for periods ranging from less than one year
to about 120 years. Within the chronosequence, autogenic succession accounts
for biogeochemical variations. (Day, Young, Porter, and Shugart)
- Above
and below ground production are limited by proximity to the fresh water
and available nitrogen. (Young and Day)
- Decomposition rate is
a function of soil moisture and thus proximity to the water table. (Day)
- Nutrients (primarily N) are derived from either N-fixation by Myrica
or atmospheric deposition and spatially are a function of exposure to oceanic
influences and soil moisture and thus proximity to the water table. (Young,
Macko and Galloway)
- Utilization of the island landscape by small
mammals varies with age and complexity of the landscape and vegetation
type. (Dueser and Porter)
- Mammalian consumers affect the composition
and structure of vegetation which, in turn alters the rates of change of
free surfaces through alteration of aeolian deposition and evapotranspiration
(Dueser, Young and Porter)
- A change in the mean level of the water
table free surface will result in a change in vegetation composition and
productivity. (Hayden, Porter, Furman, and Young)
Our model for state changes on the barrier islands is shown in 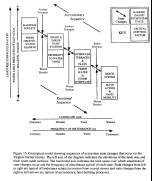 Figure 15. On these islands state changes arise from both erosional and accretionary
processes and are driven by disturbances and long-term changes in sea level.
The landscape of the northern tip of Hog Island consists of a series of
beach ridges (dunes) and swales that have built seaward over the last 120
years or so. We have dated the landforms of this landscape using survey
maps and aerial photographs in conjunction with field excavations. We have
established a chronosequence transect with permanent plots and network
of groundwater wells that record spatial and temporal variations in the
elevation of the fresh water-table. In addition, we have established productivity,
decomposition and nutrient cycling study plots along this Chronosequence
transect (Conn and Day 1993a, b). We will install large mammal exclosures
at the same locations in 1994. Chronic nitrogen-addition experiments have
been performed along the transect for the last three years. Using groundwater
wells, we are evaluating the relationships among land surface elevations,
water table free surfaces and island vegetation patterns (HI Ho-1, 2, 3,
4). In addition, we have established a water-table manipulation experiment
(see page PD 16) on isolated dunes located in a saline and hypersaline
marsh (HI Ho-1, 2, 3, 4). The fresh-water effectively "floats"
in these sand masses which are perched on a sea-water flooded, saline matrix
of fine grained marsh sediments. Hypothesis HI Ho-3 focuses the role of
disturbance in modification of the water table controlled pattern of vegetation
on barrier islands. Along the chronosequence, we are also examining the
hypotheses that proximity to the water table free surface is a principal
controller of above and below ground production (HI Ho-5), decomposition
(HI Ho-6) and nutrient availability (HI Ho-7). The utilization of landscapes
of varying age and productivity by small mammal populations will be determined
through long-term censuses coupled with structural habitat measurements
(Dueser and Shugart 1978, Dueser and Porter 1986 and Scott and Dueser 1992)
(HI Ho-8) mammal exclosures will be used to establish the effects of herbivorous
mammals on vegetation composition and structure (HI Ho-9). The water table
draw-down experiment will test the hypothesis that a change in water table
elevation will result in changes in species composition and primary productivity
(Hi Ho-10). This work is also a direct test of climate, i.e., free surface
controls on vegetation through precipitation climate change.
Figure 15. On these islands state changes arise from both erosional and accretionary
processes and are driven by disturbances and long-term changes in sea level.
The landscape of the northern tip of Hog Island consists of a series of
beach ridges (dunes) and swales that have built seaward over the last 120
years or so. We have dated the landforms of this landscape using survey
maps and aerial photographs in conjunction with field excavations. We have
established a chronosequence transect with permanent plots and network
of groundwater wells that record spatial and temporal variations in the
elevation of the fresh water-table. In addition, we have established productivity,
decomposition and nutrient cycling study plots along this Chronosequence
transect (Conn and Day 1993a, b). We will install large mammal exclosures
at the same locations in 1994. Chronic nitrogen-addition experiments have
been performed along the transect for the last three years. Using groundwater
wells, we are evaluating the relationships among land surface elevations,
water table free surfaces and island vegetation patterns (HI Ho-1, 2, 3,
4). In addition, we have established a water-table manipulation experiment
(see page PD 16) on isolated dunes located in a saline and hypersaline
marsh (HI Ho-1, 2, 3, 4). The fresh-water effectively "floats"
in these sand masses which are perched on a sea-water flooded, saline matrix
of fine grained marsh sediments. Hypothesis HI Ho-3 focuses the role of
disturbance in modification of the water table controlled pattern of vegetation
on barrier islands. Along the chronosequence, we are also examining the
hypotheses that proximity to the water table free surface is a principal
controller of above and below ground production (HI Ho-5), decomposition
(HI Ho-6) and nutrient availability (HI Ho-7). The utilization of landscapes
of varying age and productivity by small mammal populations will be determined
through long-term censuses coupled with structural habitat measurements
(Dueser and Shugart 1978, Dueser and Porter 1986 and Scott and Dueser 1992)
(HI Ho-8) mammal exclosures will be used to establish the effects of herbivorous
mammals on vegetation composition and structure (HI Ho-9). The water table
draw-down experiment will test the hypothesis that a change in water table
elevation will result in changes in species composition and primary productivity
(Hi Ho-10). This work is also a direct test of climate, i.e., free surface
controls on vegetation through precipitation climate change.
A model
of barrier island morphology (see ISLAND, SECTION 2: page PD 24), water
table, and vegetation is based on understanding the controls on the elevation
of the fresh water table free surface and depends on 1) elevation of the
land above spring tide, 2) precipitation input, and 3) evapotranspiration
losses (Rasteter 1989). Shao, Shugart, Young (1994) have developed a new
evapotranspiration model for the North Hog Island Chronosequence (see Fig.
21, page PD 24) that will be tested at our water-table manipulation experiment
site. We are also developing a maritime forest FORET model that will be
made spatially explicit by means of a water table driver in the model.
Input data to parameterize this model are being derived from a set of 100
benchmark vegetation plots on Parramore Island. At the Chronosequence Site
we are measuring decomposition, primary production, and nutrient cycling
and interpreting the observations in terms of depth to the water table
and landscape age. We are dating the landscapes of other barrier islands
of the VCR, which will permit replication sites for findings from Hog Island
studies.
The research infrastructure of the VCR North Hog Chronosequence
Site includes: 1) a photo- and map-dated 120-year chronosequence of landscapes,
2) 5 transects and mammal associated permanent plots, 3) a system of 32
minirhizotron tubes (Brown and Upchurch 1987), 4) a surveyed network of
groundwater wells, 5) a tide station, 6) an LTER minimum standard meteorological
station, 7) a solar powered experimental groundwater pumping station for
fresh-water table manipulation, and 8) permanent plots with and without
nitrogen amendments.
VCR land and lagoon free surfaces are horizontal
and parallel. The lagoon surface is a complex oscillator and the mean level
of the land relative to the lagoon surface differentiates landscapes within
the lagoon and marsh system. The central hypothesis as applied to the lagoons
and marshes site is:
The type of community and level of productivity
of emergent marsh vegetation on lagoonal marshes is controlled by the relative
elevations of the parallel marsh land free surface and the parallel, oscillatory
lagoon water free surface.
Working sub-hypotheses for Hog Island
Bay Lagoon and Marshes Site (LM) include:
- Spartina
alterniflora growth form and productivity vary as a function of the
difference between marsh surface elevation and mean sea level. Lowering
the marsh land surface will increase productivity and promote dominance
by the tall form of S. alterniflora. (Zieman and Hayden)
- VCR
Marshes that were buried during the 1933 hurricane are elevated and relatively
flat, and are therefore dominated by the short form of S. alterniflora.
(Zieman and Hayden)
- Hypersaline marshes are restricted to regions
where the marshland-surface is level at elevations between mean high tide
and spring tide. (Zieman)
- Storm-derived marine sediments deposited
below mean high tide level develop a chemistry and texture compatible with
marsh colonization and succession. (Furman and Zieman)
- Reduction
of pore water salinity by infiltration of precipitation and the activitiy
of invertebrates enhances Spartina altinaflora productivity. (Zieman
and D. Smith)
- Exchange of water between ocean and lagoon is accomplished
by a standing tidal wave and tidal currents moving through well-developed
antecedent valleys. The exchange is very efficient, and the resulting lagoonal
water body maintains a relatively constant salinity at 30-32 ppt. Exchange
is determined by water-mass trajectory path, not diffusion of stratified
flows (Oertel).
- Water mass boundaries in lagoons are determined
by differences in current "drag" created by bottom friction and
fluctuating water levels. (Oertel, Porter, and Hayden)
Our model for state changes on the lagoonal marshes of the
VCR is shown in 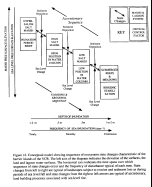 Figure 16. On these marshses state changes arise from both
erosional and accretionary processes and are driven by disturbances which
deposite large quantities of sediment and by long-term changes in sea level.
Lagoonal water and land surfaces are parallel or nearly parallel. Productivity
and height of S. alterniflora varies according to its position
in the water column (LM Ho-1). We are testing this hypothesis through our
experiment on marsh lowering and through extensive mm-resolution surveying
of marshes in the lagoons. Exceptions to parallel water and land levels
occur along the marsh edges where the land is steeply sloped and along
the marsh-island and marsh-mainland interfaces where a shallow but significant
sloping land surface is encountered. The marshes of Hog Island Bay vary
in elevation relative to mean sea level. Due to these variations the position
of marsh plants in the water column varies considerably. These spatial
variations constitute an analog of the sea level rise effects on marshes
(LM Ho-2) and to our experimental marsh lowering. The high marshes, where
evapotranspiration is high, are prone to hypersalinity. Where the marsh
slope becomes shallow, salt flats develop within a range of tidal elevations,
but if the slope is steeper, salt flats never develop, regardless of the
elevation (LM Ho-3).
Figure 16. On these marshses state changes arise from both
erosional and accretionary processes and are driven by disturbances which
deposite large quantities of sediment and by long-term changes in sea level.
Lagoonal water and land surfaces are parallel or nearly parallel. Productivity
and height of S. alterniflora varies according to its position
in the water column (LM Ho-1). We are testing this hypothesis through our
experiment on marsh lowering and through extensive mm-resolution surveying
of marshes in the lagoons. Exceptions to parallel water and land levels
occur along the marsh edges where the land is steeply sloped and along
the marsh-island and marsh-mainland interfaces where a shallow but significant
sloping land surface is encountered. The marshes of Hog Island Bay vary
in elevation relative to mean sea level. Due to these variations the position
of marsh plants in the water column varies considerably. These spatial
variations constitute an analog of the sea level rise effects on marshes
(LM Ho-2) and to our experimental marsh lowering. The high marshes, where
evapotranspiration is high, are prone to hypersalinity. Where the marsh
slope becomes shallow, salt flats develop within a range of tidal elevations,
but if the slope is steeper, salt flats never develop, regardless of the
elevation (LM Ho-3).
Along the seaward and landward margins of Hog
Island Bay, sediment supply is adequate for the formation of new marshes.
We are monitoring these new marshes to determine if sediment chemical conditions
typical of marshes develop before plant colonization or if sediment chemistry
is a consequence of marsh succession (LM Ho-4). Using remote sensing, we
have identified marshes that formed in each of the last 7 decades. These
marshes constitute a chronosequence that will permit us to use a space-for-time
substitution to detail long-term succession of marshes and to guide our
long-term monitoring of newly formed marshes. Along this chronosequence,
marsh elevations and plant productivity vary systematically as a function
of marsh age.
Preliminary studies indicate marsh primary productivity
is highly correlated with porewater salinity. Porewater salinity is reduced
directly by rainfall during low tide. Rainwater access to a thick layer
of the marsh surface depends on invertebrate burrows (macropores). We will
test the hypothesis that fiddler crabs play a primary role in regulating
primary productivity (LM Ho-5). We recognize that, in the long-term, we
will need to know a great deal more about tidal volumes, tidal velocities
and tidal residence times. Such information will be necessary to calculate
nutrient fluxes and adequately model marsh and lagoon systems. We hypothesize
that the relatively constant salinity in Hog Island Bay arises from the
lagoonal nature of the system including low fresh water input, the shallow
embayments and nearly complete tidal flushing through deep channels (LM
Ho-6 and 7)
Research infrastructure developed for the Hog Island Bay
Lagoons and Marshes Site includes: 1) a series of experimentally lowered
marsh surfaces, 2) a system of marsh surfaces with different positions
in the water column, 3) an array of marsh mm-level benchmark monuments
for surveying, 4) a GPS/Digital Fathometer System for hypsometric surveys,
and 5) a sequence of marshes of known ages for pseudo-chronological studies.
As sea level encroaches
on mainland surfaces at the VCR LTER, the transition from upland to subtidal
environments encompasses four ecosystem states: forested upland, organic
high marsh, mineral low marsh and intertidal mud flat, and subtidal lagoon
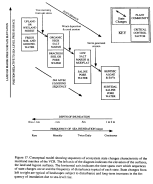 (Fig. 17). The central hypothesis stated for this landscape is:
(Fig. 17). The central hypothesis stated for this landscape is:
The vertical position of the mainland-marsh ecotone is a function
of the interaction of the non-intersecting, sloping free surfaces of the
saline lagoon water, fresh ground water table, and land surface. Within
the ecotone structured by these sloping free surfaces, multiple states
are ordered along a gradient maintained by storm-generated disturbances
against a background of chronic sea level rise. Changes in state along
the gradient are caused by mechanisms which are characteristic of each
change of state (Brinson et al. 1994).
Sub-hypotheses for the
Mainland Marsh Transition Site (MM) include:
- The forest to high marsh transition is induced by storm-generated influxes
of saline water that stresses and kills trees, tree mortality is primarily
a result of salt induced water-stress and not water-logging. (Brinson and
Christian)
- Tree mortality allows light penetration to the forest
floor and is followed by replacement of forest by marsh. (Brinson and Christian)
- The transition from organic high marsh to mineral low marsh is facilitated
by erosion of tidal creeks headward and/or along creekbank margins resulting
in subsidence of the surface of the organic high marsh. (Christian, Brinson,
Blum, and Wiberg)
- Increased oxidation of sediment organic matter
contributes to subsidence of the organic high marsh surface. Increased
oxidation is a result of better drainage created by the steeper hydraulic
gradient between the marsh water table and the creek at low tide. (Christian
and Anderson)
- The organic high marsh surface subsides in part
because of decreasing accumulation of below ground organic matter and in
part as a result of decreased allocation of resources for root and rhizome
growth. (Blum and Christian)
- Inundation and salinity by themselves
are insufficient to cause species replacements in organic high marsh communities.
Disturbance due principally to wrack deposition must be involved to maintain
within state heterogeneity (although fire, trampling, and ice scouring
may be additional disturbances). (Christian and Brinson)
- Sediment
deposition on mineral low marsh surfaces is facilitated by the frictional
resistance (i.e., baffling effects) of marsh plants and /or infiltration
of water through the marsh surface on falling tides. Erosion of marsh sediment
is inhibited by the presence of vegetation and sediment particle aggregation
by invertebrate fauna (Wiberg and D. Smith).
- Nutrient transfers
from the mainland into the mainland-marsh tidal creeks depends on the distance
of transport and the number and type of marsh zones through which the flow
must pass. (Macko, Anderson and Blum)
Our model for state changes on the mainland marshes of the VCR is shown
in Figure 17. On these marshses state changes arise from both erosional
and accretionary processes and are driven by disturbances and long-term
changes in sea level resulting in increased frequency of inundation of
uplands by saline waters.Work at the Mainland Marsh Site has focused on
the position of marshes in the coastal landscape as a function of the recent
history of rising sea level and existing upland geomorphology (see Fig.
3b, page RPS 3). We have simplified the mainland-marsh-tidal creek landscape
to 4 states (Fig. 17), and have proposed mechanisms responsible for state
changes that could be expected as a result of sea-level rise (Brinson et
al., 1994). While our model for the changes resulting from sea level rise
has some similarity to that of Dame et al. (1992) our focus is mostly on
the marsh surfaces rather than the tidal creek changes. Our previous work
in assessing these state changes (and maintenance of the states) has been
both observational and experimental. Most of the observational studies
are long-term and ongoing at the Phillips Creek marsh. This will remain
our reference site for mainland-marsh ecotone state changes and dynamics,
while Mill Creek marsh will be the focus of our proposed long-term experimental
simulation of tidal creek headward erosion. In the organic high marsh,
we track the movement of the interface between patches of Juncus roemerianus
and its neighboring plant species (short-form Spartina alterniflora
and S. patens/Distichlis spicata association) at different
locations reflecting different states and flooding regimes. Further, we
examine aspects of both above and below ground dynamics inside and outside
these patches (Blum, 1994). We also use aerial photographs to examine transitions
from forest to marsh and marsh to mud flat (Kastler 1993 and Fig. 3b, page
RPS 3), and will use similar techniques to monitor tree mortality as a
result of storm-induced upland flooding (MM Ho-1) and to track headward
erosion of tidal creeks (MM Ho-3).
An experiment that increases
inundation frequency by pumping creek water onto marsh plots was initiated
in summer 1993 (see Section 2, Mainland Marsh Inundation Experiment, page
PD 18). This experiment examines the role of inundation and wrack deposition,
separately and in combination, in maintaining organic high marsh plant
diversity in Juncus roemerianus dominated and Spartina patens/Distichlis
spicata co-dominated associations (MM Ho-6). Primary productivity,
species displacement, sediment chemistry (primarily salinity, redox potential,
and species of N and S), and trace gas fluxes (CH4, CO2, N2, N2O) are monitored during these
pumping experiments (MM Ho-8). We propose to begin experiments during winter/spring
1995 (seasons of peak storm frequency and intensity) in salt loading and
tree girdling to observe 1) thresholds of salt effects on stress symptoms
and mortality (MM Ho-1) and 2) response of the ground cover to increased
light penetration (MM Ho-2). Experiments to address MM Ho-3, 4,
5, 6, and 7 will be done in a long-term manipulation experiment in another
marsh as described in Section 2 of the Project Description: Marsh Mainland
Inundation Experiments.
Other proposed studies include determination
of 1) the primary sediment transport mechanisms from marsh surfaces, both
depositional and erosional (MM Ho-7), 2) the role of the biota in modifying
sediment transport by altering marsh surface frictional resistance and
particle aggregation (MM Ho-7), and 3) the movement of agricultural chemicals
in groundwater to forested buffer strips, marshes, and tidal creeks (MM
Ho-8). Hypotheses concerning mechanisms of sediment deposition onto and
erosion from marsh surfaces will be tested on a mineral low marsh located
proximally to tidal channel and lagoon sediment sources and to faster moving
water in tidal creeks and channels. Effects of plant stem density and marsh
elevation on sedimentation will be investigated through measurement of
flow velocity, suspended sediments, and marsh elevation on natural and
lowered marsh surfaces (see Section 2: Long-Term Experimentrs, page PD
16). In addition, we will begin studies to examine the relationship of
marsh surface invertebrate fauna abundance, biomass, and species composition
(Yozzo et al. 1994) to the intensity of sediment aggregation and aggregate
properties (size, settling velocity, and durability) (Kastler 1993).
Stable isotope studies to characterize nutrient transfer across the land,
fresh-water table, and sea-level free surface interfaces were begun in
1993 at the Mainland Marsh Site. The site is ideal for this type of study
because large quantities of fertilizers are applied to the highly permeable
upland soils of this area. Isotopic fractionation of nitrogen and carbon
species and flux measurements of CO2, CH4, N2O, and NOxwill be used to identify biogeochemical
processes that lead to losses of inorganic nitrogen and carbon (Macko et
al. 1987) due to anthropogenic disturbances (Anderson and Levine 1987)
(MM Ho-8). We will also measure rates of nitrification and denitrification
using an 15N dilution technique to complement our stable isotope
measurements (as non-LTER funds become available).
Research infrastructure
of the Mainland Marsh Transition Site includes: 1) transects across the
marsh and into the neighboring forest of known elevation, distance from
tidal water source, flooding regime, and related physical and chemical
variables, 2) a network of water-level recorders and tide gages, 3) a solar-powered
meteorological station , 4) experimental plots augmented with lagoon water
using a solar-powered pumping system, 5) a network of ground water wells,
and (6) a series of permanent plots to monitor changes in the interface
between J. roemerianus and neighboring marsh plant associations.
PROJECT DESCRIPTION
VCR long-term experiments
consist of manipulations of critical free surfaces: land level, sea level
and fresh water-table level. Elevations of these free surfaces are critical
state variables in ecosystem dynamics. Our general hypothesis is that changes
in elevation will bring about transitions to new states that are either
self-maintaining or successional. We propose long-term low-level manipulations
of free surfaces rather than singular disturbance and recovery experiments.We
will alter tidal inundation in marshes using 3 approaches to test different
hypotheses and the level of the fresh water table on a section of a barrier
island. Each experiment will be paired with a spectrum of natural analogs
that have experienced variation in that critical free surface in the last
several decades. In addition, we will use the results of the experiments
and observations from the natural analogs to parameterize a model of each
system. The measurements to be taken in each experiment are summarized
in Table 3.
The terrestrial surface of the VCR is a mosaic of discrete patches, with
abrupt edges between communities dominated by different plant assemblages.
A well developed dune-swale unit exhibits segregated communities (i.e.,
grass-dominated ridge vs. shrub-dominated swale) related to elevation of
the water table. Young (1992) has demonstrated that the dominant shrub
on the islands, Myrica cerifera, occurs in close proximity
to ground water reserves (i.e., swales) because of drought sensitivity.
Our proposed experiment to reduce the size of the confined fresh water
lens by pumping should lead to drought stress, reduced production, and
increased mortality. By comparison, experimental addition of water to the
more xeric grass-dominated dunes should enhance water availability and
lead to successful establishment of Myrica and other woody species.
The edge or ecotone between the two associations will change through time
in response to the manipulation of the fresh water lens.
Parramore 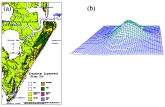 (Fig.
18a) and Hog Islands have round sand masses (Fig. 18b) with isolated freshwater
reserves perched atop sand flats and surrounded by saline and hypersaline
salt marshes. In 1993, we proved we can control the level of the water
table in these pimples by pumping. We have selected three similar pimples
(Fig.
18a) and Hog Islands have round sand masses (Fig. 18b) with isolated freshwater
reserves perched atop sand flats and surrounded by saline and hypersaline
salt marshes. In 1993, we proved we can control the level of the water
table in these pimples by pumping. We have selected three similar pimples
 (Fig. 18c on next page) for our initial draw down experiments in 1994.
One will be a draw-down treatment in which the water table is lowered at
a rate equal to an increase of 1 mm d-1 of evapotranspiration.
A second pimple will receive the removed water as a supplement equivalent
to an increase in rainfall of 1 mm d-1 added to the system
below the surface of the sand. The third pimple is our control. In 1994
will will add wells, pumps and recorders to two additional replicates of
each treatment and control. In 1995 two more replicates will be installed
and bringing the total sample size to five. We expect to continue these
experiments for at least the next decade. Pumping studies in 1993 did not
involve a manipulation of the water table (Fig. 18d). Pumped water was
re-injected below ground at the same location. We established that we can
pump at a rate of 1 to 3 mm d-1 and can record mm level changes
in the water table. We also demonstrated that we can record community level
evapotranspiration within each day and can record the seasonal draw down
and recharging of the water table (see Fig. 4b, page RPS 4).
(Fig. 18c on next page) for our initial draw down experiments in 1994.
One will be a draw-down treatment in which the water table is lowered at
a rate equal to an increase of 1 mm d-1 of evapotranspiration.
A second pimple will receive the removed water as a supplement equivalent
to an increase in rainfall of 1 mm d-1 added to the system
below the surface of the sand. The third pimple is our control. In 1994
will will add wells, pumps and recorders to two additional replicates of
each treatment and control. In 1995 two more replicates will be installed
and bringing the total sample size to five. We expect to continue these
experiments for at least the next decade. Pumping studies in 1993 did not
involve a manipulation of the water table (Fig. 18d). Pumped water was
re-injected below ground at the same location. We established that we can
pump at a rate of 1 to 3 mm d-1 and can record mm level changes
in the water table. We also demonstrated that we can record community level
evapotranspiration within each day and can record the seasonal draw down
and recharging of the water table (see Fig. 4b, page RPS 4).
Experimental
hypotheses include: 1) increases/decreases in the elevation of the water
table free surface in the pimples will result in reduced/increased grass
cover on the pimples, increased/reduced woody plant establishment and growth,
expansion/contraction of the Iva and Baccharis shrub, and
Spartina patens grass zone around the pimples at the expense of
the surrounding salt marsh, 2) fluctuations in the elevation of the water
table free surface will lead to significant variations in nutrient budgets,
especially nitrogen input via Myrica N-fixation, and 3) differential
responses to changes in water table free surface will lead to shifts in
species composition within the pimple community. Over the long-term, these
experiments will constitute a climate change experiment. We anticipate
that the pimple from which water is extracted will experience the equivalent
of a 25% rainfall reduction, the receiving pimple a 25% effective increase
in average annual rainfall. The control pimple will have no manipulation
of the water table free surface other than that arises from year to year
variation in rainfall.
Analog sites to the experiment. Depth to the
fresh water table free surface depends on the elevation of the pimple surface.
In that the pimples are quite variable in elevation, they differ a great
deal in the water resource available to terrestrial vegetation and, as
such, the vegetation cover across the spectrum of pimples includes several
biome-level vegetation types, e.g., open dunes, grasslands and forests.
The smallest, lowest elevation pimples are entirely S. patens grass
covered systems. Pimples with slightly higher elevation are entirely covered
with shrubs (Iva and Baccharis or Myrica). Larger
pimples have mature Juniperus virginica and Pinus taeda forests
with a grassland/desert center in the pimple. Using surface wells, we will
initiate water table monitoring of dozens of pimples in order to determine
community wide evapotranspiration rates from different communities and
the role of rainfall variation on pimple community structure over time.
Measurements of 18O content will be used to extimate in water
turnover rates in the pimples. These isolated freshwater systems will permit
us to study biome level evapotranspiration differences in a common climate
regime. Comparisons of pimples with and without Myrica aided by
15N measurements will facilitate the assessment of the significance
of Myrica nitrogen-fixation to the nitrogen budget. We will use
the data from these monitoring programs to further parameterize our evapotranspiration
model (see Section 2: Models and Synthesis page PD 23).
In 1992 we began
a program of experimental manipulation of the land free surface elevation
along the margins of salt marshes to artificially bring about changes in
the position of the marsh in the tidal water column. This experiment is
prompted by two observations. First, compared with other marshes in the
region, the lagoon marshes of the VCR seem to be higher and flatter (LM
Ho-2), and less productive, and second, when large blocks of these elevated
marshes are undercut by natural erosion forces and drop 10-20 cm onto the
creekbank, the plant biomass and productivity greatly increase. From this
we hypothesize that modest accelerated sea level rises will result in increased
Spartina alterniflora biomass and productivity. Preliminary tests
indicate that the height and standing biomass of the Spartina increased
even in the first year (LM Ho-1).
The experiment involves undercutting a large block of creekbank marsh
 (Fig. 19a) and severing its attachment
at the sides and back so it remains in place laterally, but is lower in
the water column (LM Ho-1). This will allow increased flushing by tidal
water. In 1992, we used hoe-like implements to remove sediment from beneath
the root zone and hand saws to sever the connections. This procedure was
very difficult and often yielded an uneven surface. In 1994, we will use
a hydraulic pressure washer to remove sediment from beneath the root zone.
The back and edges will then be severed with a pressure cutting head (3500
psi) and a hand saw, and the block allowed to settle into place. The result
is a lowering of the marsh surface in the water column of approximately
5 cm.
(Fig. 19a) and severing its attachment
at the sides and back so it remains in place laterally, but is lower in
the water column (LM Ho-1). This will allow increased flushing by tidal
water. In 1992, we used hoe-like implements to remove sediment from beneath
the root zone and hand saws to sever the connections. This procedure was
very difficult and often yielded an uneven surface. In 1994, we will use
a hydraulic pressure washer to remove sediment from beneath the root zone.
The back and edges will then be severed with a pressure cutting head (3500
psi) and a hand saw, and the block allowed to settle into place. The result
is a lowering of the marsh surface in the water column of approximately
5 cm.
In both years of the experiment to date a 3- to 5-fold increase
in plant biomass in the lowered plots compared to control sites on the
mainland and island marshes was observed. In the first year, the primary
source of biomass increase at the mainland sites was stem density, while
it was largely plant height on the island marshes (Fig. 19a). In the second
year both factors had increased at all sites.
Two adjustment scenarios
are planned: acute and chronic. In the acute scenario, the marsh will be
lowered by 15 cm creating a marsh surface equivalent to the 1930s marshes.
In the chronic scenario, the marsh will be lowered about 5 mm every other
year, thus approximating the current rise in sea level at the VCR. Above
ground marsh productivity (Morris et al. 1990) and peak standing crop will
be monitored yearly and total biomass (above and below ground) will be
measured every second year. We hypothesize that secondary production will
also increase with increasing relative sea level. Specifically, we expect
increases in juvenile fish and crustacean utilization of the treated sites.
Sedimentation and marsh elevation will be carefully monitored on test and
control plots and local marshes.
Marsh manipulations will be performed
along the margins of mid-lagoon marshes. Each manipulation will result
in the lowering of 4 m2 of marsh surface and will be paired
with an adjacent control surface the same size. Both treatment and controls
will have 5 replicates. To assess the possibility that the experiment records
simple edge effects we will also survey marshes in Hog Island Bay with
different relative elevations and test the hypothesis that relative marsh
elevation results in the same effects as in the experiments.
Throughout
the VCR there are hundreds of natural examples of the creation of high
productivity low marsh habitat from the erosion and slumping of the perched
short-form Spartina marshes. Many of these do not survive because
they do not readily develop a prograding marsh face and are rapidly eroded
by wave action in the tidal creeks. Where such an interface develops the
new tall-form marshes can last many years. We will document these slumps
on our GIS system, label the sites, and follow them on an annual basis
to track their long-term fate. In addition, because marsh burial in the
1930s varied considerably in depth of burial, present-day marshes vary
in elevation. We are surveying marsh elevations at the mm level in order
to identify a natural gradient in marsh submergence that will serve as
an alternate source of information equivalent of that to be experimentally
determined.
As sea level rises and the frequency and intensity of disturbances
increase, a sequence of state changes occurs in the mainland-marsh ecotone
(Fig.17, page PD 13). We propose experiments to simulate headward erosion
of tidal creeks that occurs as a result of sea-level rise (MM Ho-3, 4 and
5) and to simulate the effect of increased inundation frequency and disturbance
(specifically, wrack deposition) in the organic high marsh (MM Ho-6).
Inundation Experiment: In our experiment to test MM Ho-6, currently in
place at Phillis Creek, we alter inundation by pumping water into experimental
plots with a battery-operated submersible pump charged by solar power.
The pumping occurs when water in the creek rises above a designated water
stage. The water is distributed throughout 3 x 4 m wood-framed plots which
allow ponding on the surface during pumping followed by slow infiltration
and exit of the flood water. It takes approximately 45 min. to cover a
plot with water. Inundation plots are positioned so that each contains
two different plant associations (e.g., 2 x 3 m in a Juncus roemerianus
patch and the other 2 x 3 m is in an area of Spartina patens and
Distichlis spicata). Pore water samplers (sippers) to assess sediment
chemistry are located inside and outside of the plots (Table 3). Further,
half of the plot stretching across both plant associations is covered with
Spartina alterniflora wrack. We are using a randomized block design
of 3 blocks each with 3 plots. Each block consists of one plot with increased
inundation and two control plots 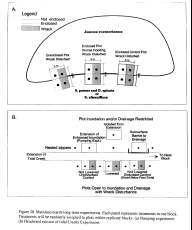 (Fig. 20a). One block is represented in
Fig.20a. A prototype installations were completed during 1993. The additional
blocks, each located in the organic high marsh, have been established and
are currently fitted for 1994 when inundation frequency will be increased
and the plots disturbed by additon of wrack. This design tests disturbance
effects from wrack deposition with and without associated inundation increase
(MM Ho-6). We will include further testing of this hypothesis at the Mill
Creek Experimental marsh as part of the tidal creek headward erosion simulation
experiment described below.
(Fig. 20a). One block is represented in
Fig.20a. A prototype installations were completed during 1993. The additional
blocks, each located in the organic high marsh, have been established and
are currently fitted for 1994 when inundation frequency will be increased
and the plots disturbed by additon of wrack. This design tests disturbance
effects from wrack deposition with and without associated inundation increase
(MM Ho-6). We will include further testing of this hypothesis at the Mill
Creek Experimental marsh as part of the tidal creek headward erosion simulation
experiment described below.
Tidal Creek Headward Erosion Simulation:
Mill Creek marsh, like the Phillips Creek marsh, is in the Machipongo drainage
system and is located landward of Fowling Point marsh. In addition, Phillips
and Mill Creeks are connected by the Eastern Lead, an important tidal channel.
The Mill Creek marsh site selected for manipulation is approximately 8
ha, bordered on two sides by small sandy islands or "necks",
partially blocked at one end by a causeway, and connected to Mill Creek
via a short tributary. A randomized block design will be used of at least
5 blocks with elevation and disturbance nested within exposure to increased
tidal flooding will be used. One block is represented in Figure 20b. The two plots adjacent to the extension will receive
enhanced flooding in contrast to the third "isolated" plot (MM
Ho-3, 5). One of these two plots will have a plywood barrier embedded into
the soil near the extension to restrict drainage such that comparison with
the other "adjacent" plot tests MM ho-4.
The plots below
the extension in Fig. 20b provide alternate testing of MM Ho-3, 5, and
6. the design is similar to that used in the pumping experiment with disturbed
and undisturbed controls, a plot with enhanced inundation (this time by
lowering the marsh surface as previously described), and nesting within
each treatment for the influence of wrack deposition. The lowering will
enhance inundation beyond that achieved by extending the creek alone. Plots
will be 2 m long by 3 m wide, oriented with the widest portion of the plot
parallel to the ditch (Fig. 20b). With this method of simulating sea-level
rise there is less control of hydrology than with pumping but greater ability
to mimic sediment transport and rising sea-level effects in upper tidal
creeks. This will allow us to directly address MM Ho-3 through 7 by monitoring
changes in the biological, chemical, and physical characteristics of the
manipulated areas (Table 3).
Connection to modeling: Part of our effort
on marshes is to identify and quantitatively model depositional and erosional
mechanisms on marshes in response to tidal and storm-induced flows (Wiberg).
We have also developed a mechanistic model of salt marsh organic matter
accumulation based on plant production and decay and constrained by sediment
physical/chemical properties (Nuttle and Blum). Results from the proposed
experiments and continuation of experiments at the Mainland Marsh Site
will allow us to test the validity of our model for differing sediment
properties that are determined by differences in tidal inundation frequencies.
In the long-term, we propose to link the sediment transport and organic
matter accumulation models to allow us to predict patterns of marsh accretion
and loss (Nuttle, Wiberg, Blum, Brinson, and Christian).
The holdings of the VCR long term data sets are presented in Table 4a&b.
THE FIVE CORE AREAS
The five core
areas provide a common currency with which to compare the four main VCR
LTER research sites, as well as other LTERs and LMERs. Additionally, the
core-area work provides a long-term set of reference measures for comparison
with our long- and short-term experiments. At each of the four research
sites, we measure primary production, organic matter accumulation, and
trophic structure within the context of large transient and small progressive
changes in the position of the the free surfaces that result from disturbance,
the fifth core area. The core area measurements we make are dictated by
the characteristics of the site and the hypotheses being addressed at each
of the sites and thus are not necessarily the same at all research sites
since the variety of states at the VCR is so diverse. As a preamble to
the discussion of our work in the five core areas, an overview of activities
is provided in Table 5 and Table 6 lists the hypotheses that directly appply
to core area measurements.
Primary production on the VCR Megasite is assessed on large spatial
and temporal scales using remote sensing techniques. Aerial photography
and satellite imagery are used to examine landscape-level vegetation responses
to changes in the intersections of the three free surfaces over time. With
our library of high resolution aerial photographs, we can map important
landscape elements back more than 45 years. Approximations of long-term
net production are based on differences between area-based biomass estimates
for various land-cover types. In order to use remote sensing to "scale
up" site-based studies of primary production, we are collaborating with
NASA on the use of advanced satellite and airborne scanners (TM, AVIRIS,
and MODIS) to estimate primary production over wide areas for short time
frames. MODIS, AVIRIS, and TM data, along with data from a ground based
sun photometer (for atmospheric correction) were collected as part of the
SCAR-A campaign during the summer of 1993. We intend to extend this collaboration
in a joint LTER/NASA proposal aimed at calibrating MODIS-based remote sensing
techniques for measuring land cover, primary production and leaf-area index.
At the North Hog Island Chronosequence Site, we have established permanent
vegetation plots across the dune-swale chronosequence where the dominant
plants are Myrica and Ammophila, a keystone species (see
Relationships, page PD 6). We measure above ground biomass annually as
peak standing stock and leaf area index in the Myrica thickets.
Above ground production Myrica is measured as a relative index based
on number and length of new shoots produced each year and root turnover
is measured using mini-rhizotrons for all plant associations along the
chronosequence. These measures focus on the relationship of primary production
to dune age, depth to the fresh water table and nitrogen availability (see
page PD 21).
The approach used for the Hog Island Bay Marshes and Lagoon
Site includes 1) a series of permanent Spartina alterniflora (also
a keystone species) plots in recently deposited substrate (< 20 years)
and older sediments (> 60 years, but estimated to be > several hundred
years) and 2) a transect of permanent water column stations from mainland
tidal marsh creeks, across the lagoon, in barrier island marsh creeks,
to an ocean inlet. The positions of the marsh vegetation plots are stratified
based on the relative position of the land and sea water free surfaces
and include high and low marsh plots. Above ground production in the lagoon
and barrier island marshes is measured using the method of Morris and Haskin
(1990). Peak standing stock is also measured in four marshes in the lagoon.
Along the water column transect, phytoplankton standing stock is measured
monthly as chlorophyll-a concentration. Beginning in summer 1994, we plan
to measure phytoplankton production on a quarterly basis using radiolabeled
bicarbonate incorporation (Malone 1982).
In the mainland marshes, we
have established a series of permanent vegetation plots in the organic
high marsh and mineral low marsh where we measure both above and below
ground plant production to study the role of disturbance in maintainance
of plant diversity and the relationship between frequency of tidal inundation
and allocation of biomass to above and below ground portions of the plants.
These plots include the keystone species, Spartina alterniflora,
as well as Spartina patens, Distichlis spicata, and Juncus
roemerianus. For above ground production of S. alterniflora
we use the Morris and Haskin (1990) method, of J. roemerianus the
method of Christian et al. (1990), and of S. patens/D. spicata
association a sequential clip plot method. For below ground production
we use the method described in Blum (1993).
At the Megasite scale, we have completed a
synoptic survey of the percent organic matter in forested barrier island
soils, in sediments of Hog Island Bay, and in barrier island fringe, lagoon,
and mainland marshes. We will repeat this survey every 5 years: the next
survey is scheduled for fall 1997. To understand the factors controlling
organic matter accumulation, we measure 1) below ground production and
decay at the Hog Island Bay Marsh/Lagoon and Mainland Marsh Transition
Sites as described in Blum (1993); and at the North Hog Island Site, 2)
root turnover and decay as described in Day (1993) and Weber and Day (1993)
and 3) leaf litter fall and decay ( by M. Harmon, Andrews LTER LIDET program).
Additionally, at the Mainland Marsh Site we are working to determine the
relative contribution of new vs. old organic matter to sediment organic
matter accumulation by comparing organic matter in plots where organic
matter produciton is prevented in comparison with untreated, adjacent plots.
We have been using wooden dowels to measure relative above and below
ground decay rates at three marsh locations for three years. We will expand
this effort to all of our main research sites and 3 long-term experiments
during the summer of 1994. Through the use of this technique we will be
able to compare relative decay rates among locations at the VCR LTER as
well as with those LTER\qs and other long-term experiment projects that
are participating in the LIDET study (see Intersite Activities, page PD
27).
Nutrient movement among landscape elements at the VCR occurs via groundwater
transport and general circulation of lagoon water. To understand nutrient
transport at the Megasite scale, we are examining the exchange of water
between the ocean and the lagoon and characterizing the lagoon\qs hypsometery.
Also, we have established a transect across Hog Island Bay where we measure
N and P standing stocks (as well as other chemical/physical parameters)
at 10 permanent stations. To understand the effect of disturbance and
constraints on both primary production and organic matter accumulation,
we monitor nutrient standing stocks of N and P in all of our primary production
and organic matter accumulation plots at three of the main research sites
(not the Megasite). Nitrogen pools and fluxes in both sediments, porewater,
and plans are measured on the Lagoons and Marshes Site. We also measure
salinity in a series of groundwater wells at the North Hog and Mainland
Marsh research sites because of our concern with the relationship between
the fresh-water and sea-water free surfaces. Many of our sites have anaerobic
substrates which have a signficant impact on nutrient transformations;
thus, we also monitor indicators of anaerobiosis such as dissolved oxygen
and sulfide concentrations.
Nitrogen cycling is especially important
at the North Hog Island Chronosequence and the Mainland Marsh Transition
sites. Because plants along the North Hog Island Chronosequence are nitrogen
limited (Day 1993, Weber and Day 1993), we measure N mineralization rates,
symbiotic dinitrogen-fixation by Myrica, and collect data on atmospheric
inputs of nitrogen in addition to standing stock measures. Mainland marsh
sediments have elevated nitrogen concentrations with respect to lagoonal
and island marshes. These differences may reflect the influence of agricultural
practices on adjacent uplands or differences in the source of marsh sediments
(i.e., flooded upland soils vs. recent oceanic deposits). We use a system
of groundwater wells at the Mainland Marsh Site to monitor stable N isotope
composition of upland groundwater, sediment pore water, and tidal creek
water to elucidate sources of nitrogen input into the tidal creeks and
lagoon. Because of the importance of marshes in nitrogen cycling, we have
been measuring nitrogen mineralization, denitrification, nitrification,
and dissimilatory nitrate reduction using 15N-pool dilution
techniques. Additionally, we will begin measurement of nitrogen gas fluxes
to quantify N losses from the marsh.
The complex juxtaposition of free surfaces which characterize
the VCR Megasite provides a unique opportunity to test theories which contrast
the roles of isolation, habitat sufficiency and population processes in
determining the composition of island faunas. In partnership with the Virginia
Museum of Natural History, allozyme analyses of small mammals collected
during multi-island surveys are being used to examine the effects of isolation
on the genetic diversity of populations to attempt to reconstruct the colonization
history of the islands and to identify population sources for the island
populations. In Hog Island Bay, we are using stable isotopes to trace aquatic
foodwebs and are comparing the VCR foodwebs with those of nearby Chesapeake
Bay. At the scale of the North Hog Island Chronosequence we examine the
relationship between the complexity and age of the vegetation and small
mammal (consumer) populations using long-term census techniques coupled
with structural habitat measures. At the Hog Island Bay Marsh Lagoon and
Mainland Marsh Transition Sites we examine marsh surface utilization by
sub-adult and adult nekton and the role of invertebrate burrowing activities
on primary production. Total nekton abundance, microhabitat selection,
size-frequency distribution, and species composition of resident and non-resident
nekton are compared in regularly and irregularly flooded marsh areas. Preliminary
work to examine the effect of fiddler crab burrow density on S. alterniflora
production was done during the past summer. Similar experiments will be
done in conjuction with the Tidal Creek Headward Erosion experiment.
Recently (Spring 1993) we began studies to determine the contribution of
microorganisms to higher trophic levels and have been developing techniques
(Garland and Mills 1991) to assess functional and structural differences
in the microbial community of mainland marsh tidal creeks. We plan to initiate
studies as part of the North Hog Island Chronosequence, in which we establish
exclosures to assess the effects of herbivory on the compostion and structure
of insular vegetation.
Understanding the role of disturbance in system dynamics is a critical
part of our research program. It is our view that disturbance 1) is essential
to the maintenance of certain system states, 2) regulates succession, 3)
changes system states, and 4) is responsible for the VCR\qs diverse landscape.
Disturbance occurs mainly in the form of coastal-storm related phenomena.
Regular, predictable fluctuation of the sea water free surface is commonplace
throughout the system, and includes semi-diurnal tidal flooding, lower
frequency flooding due to astronomical highs in the monthly tide cycles,
and longer-term, more monotonic changes in sea level. Superposed on these
patterns are the aperiodic disturbances such as storm surge flooding, sediment
and wrack depostion, erosion, and fires. Although tropical storms are
infrequent -- several landfalls at the VCR per century since 1650 -- the
magnitude of their disturbance is great. Overwash events move tremendous
volumes of sand and salt water resulting in saline intrusions into fresh
water stores of the islands and mainland. We have installed a system of
measurement stations at the VCR Megasite scale that, when coupled with
appropriate aerial and satellite images, permits long-term monitoring of
both regular relatively high frequency fluctuations, and aperiodic, lower
frequency disturbances. In addition to continued monitoring disturbance
events, we collect supplemental information at the North Hog Island Chronosequence,
Hog Island Bay Marsh and Lagoon, and Mainland Marsh Transition Sites to
determine the extent of disturbance (erosion, shore-line change, salinity
changes, etc.) and to establish new baselines.
VCR modeling efforts brings a diverse array of modelers
and modeling approaches includeing state variable change models, disturbance
frequency models, material flux model, and sediment dynamics models, to
bear on the central problem of understanding state change and dynamics
on the VCR LTER.
Individual-based models (Huston et al. 1988) have been developed to simulate
succession to a quasi-equilibrium or ecosystem steady state for the VCR
and Luquillo LTERs, respectively, Bonan and Hayden 1990a, b; O'Brien et
al. 1992) and are being developed for a network of LTER sites (Shugart
et al. 1991 a 1992 a; Smith et al. 1992 a, b). A new model of this type
will be developed for the island pine forests of the VCR and parameterized
with data collected in 1993 for 100 permanent plots on Parramore Island.
This model will emphasize water balance and will be controlled, in part,
by the "natural lysimeter" aspect of the VCR islands land and
water table free surfaces and will be tuned using data collected in our
water table draw down experiment (see Long-term Experiments page PD 16).
Unlike most terrestrial systems, the VCR islands are isolated, almost hydroponic
systems. The fresh water reserves are perched on a saline, sand matrix
limited horizontally by the ocean and the lagoon. Fresh water input is
only from rainfall and little of the rainwater is lost as runoff because
of the high hydraulic conductivity of the soils. The size of the reservoir
is determined by island topography (Strack 1971; Noest 1991; Bolyard et
al. 1979). Loss of fresh water is predominantly by evapotranspiration (ET)
and evaporation from small ponds. Maximum LAI and life form vary with rainfall
input and island topography (Eagleson 1982, Woodward 1987). We have developed
an ET model for barrier island vegetation (Shao et al. 1994
 (Fig. 21).
(Fig. 21).
We will develop a FORET type model (Shugart 1984) of island vegetation
"perched" on a barrier island "lysimeter" and parameterized
using draw-down experiment data. The model will be used to analyze ecosystem
changes due to coastal storms and overwash of sand (changes in island elevation)
and to changes in island width (shoreline erosion due to storms and shoreline
erosion due to sea level rise). The proposed model will permit direct inclusion
of critical climate change variables (storm frequency and sea level rise)
into the FORET model genre of models. Because models of this type have
been developed or are under development for all forested LTER sites (often
in cooperation with the modeling group at the VCR LTER), the development
of this model will also promote LTER intersite comparisons.
A key tenet of the
VCR research program is that when an ecosystem state variable(s) exceeds
a threshold, the state of the system and the ends of succession are fundamentally
changed (Hayden et al. 1991). We further envision that both geophysical
and biological state variables, when thresholds are exceeded, give rise
to a state change. In the ISLAND model developed for Hog Island by Rastetter
(1991) a species extinction at a specified time in model years can give
rise to a new island state. Rastetter's model ISLAND is a Markov model
of vegetation on cell based transects across an island with a specified
topography controlled by ocean delivery of sand and trapping of sand by
plants (see also Noest 1991). These state variable change models will be
parameterized with data from our Megasite surveys and form data collected
at the Chronosequence Site.
We will determine temporal transition probabilities
for ISLAND using a series of aerial photographs, collected at ~5 year intervals
and with a spatial resolution of 2 m. This data base will permit an estimate
of the spatial variability of transition probabilities of ISLAND on a transect
specific basis (see Fahrig et al. 1993b). The geophysical details to run
ISLAND are available at 50 m intervals along Hog Island. The output of
ISLAND, when parameterized in this way, will be interfaced through ARCINFO
to make ISLAND a spatially explicit, three dimensional landscape model.
This will allow us to test system state changes due to extinction and invasion
at local and island-wide spatial scales. In addition, geophysical state
changes arising from changes in shoreline erosion, storm frequency, overwash
frequency and relative sea-level rise rate changes can be studied using
a common dynamic structure.
We will also develop an ISLANDII model of
the dynamic interactions of grasses, shrubs and trees in an individual-based
simulator framework. This model has implications for a variety of mixed-life-form
systems (see Wu et al. 1985; Sharpe et al. 1985, 1986 for related approach).
Under other funding, one of us (Shugart) is involved directly with an effort
to compare similarly derived models at several LTER sites. We have also
been designated as coordinators of an international effort to develop mixed-life
form simulators (such as the modified ISLANDII model) for savanna ecosystems.
The modeling group (Jean-Claude Menaut, France and the Ivory Coast; Ian
Noble, Dean Graetz, and Brian Walker, Australia; Oswaldo Sala, Argentina;
Peter Frost, Zimbabwe; Bob Skole, South Africa) met at the VCR LTER in
May 1992 to initiate the development of a shared mixed-life-form model
for global applications. Our most likely role will be to lead in water/plant
interaction simulation, project coordination, and development of the central
competition program. Our contribution will pivot on the ISLANDII model.
A salt marsh model for use
at the Lagoons and Marshes and mainland Marshes Sites is being developed
to examine the effects of sea level rise and differential sedimentation
on the growth, biomass, and species succession on the VCR salt marshes.
The basis of this model is a refined version of the MARSH model developed
by Zieman et al. (1977) for predicting salt marsh succession on natural
and created marsh sites. Plant growth and succession are controlled primarily
by physical variables such as tidal height and range, salinity, temperature,
and light. These variables mirror the control postulated to dominate in
such situations. Our sampling of long-term plots for biomass and productivity
will serve to refine the parameters for local marshes.The model now simulates
a series of points along a transect from the mudflats to the uppermost
edge of the marsh. Future refinements will include the addition of nutrient
dynamics and linkages to the sediment deposition models described below.
For the
Chronosequence Site, Fahrig (1990a, b) determined that Markov transition
probabilities for occurrence of Hog Island plants were strongly dependent
on: (1) their proximity to and variability of the shoreline and (2) variability
in the proximity to the inland limit of penetration of overwashed beach
sand. These measures of island variability are available at 50 m intervals
along the 8 km of Hog Island. Additionally, we know that the variations
in the position of the shoreline and the overwash penetration limit are
related to the frequency of coastal storms for which we have a 100 year
record (Hayden 1981). Recently, we have written a statistical simulation
model for both shoreline variability and overwash penetration limit variability.
With these models defining the quasi-equilibrium of island geomorphology
for the period 1941-1977 and using Fahrig's transition probabilities for
the same period, we propose to use the model to produce the null statistics
against which the observed state changes since 1977 period can be compared
and used as a model test. We also used Fahrig's model to compare the disturbance
response of vegetation at the VCR LTER with that at the CPR LTER (Fahrig
et al. 1993).
Below ground organic matter is particularly important in marshes because
it is the largest component of net primary production (based on root/shoot
measurements, Gallagher 1974; Stroud 1976; Good and Frasco 1979; Valiela
et al. 1976; Stout 1978) and because it has a volume equivalent to that
of the mineral matter in most marsh soils (DeLaune and Patrick 1980). Organic
matter is also important to soil diagenesis (Howarth and Teal 1980; Teal
1962), response to sea level change (DeLaune and Patrick 1980), and succession
in marshes (Gray and Bunce 1972). Because organic matter dynamics are so
important in marsh systems, we developed a marsh organic matter model as
a tool for synthesis and a guide for determining research priorities in
our marsh research. The model focuses on net primary production, turnover,
and decomposition of structural material (i.e., the materials with the
greatest potential for long term storage as sedimentary organic matter).
We specifically focused on developing a model to estimate the sources of
bias in estimation of net below ground production and in interactions between
timing of root growth, translocation of photosynthate, and decomposition
of dead material. The model simulates production/turnover and decomposition
dynamics of structural carbon in marshes, and ignores other critical but
difficult to estimate aspects of the carbon budget such as respiration
and root exudates. We hypothesized that these fluxes impact less directly
on dynamics of the structural pool of carbon. This provided reasonable
simplification of model structure necessary to proceed. The model has served
as a data source (based on South Carolina and Georgia data for parameterization)
for investigating sensitivities of harvest method to sample frequency and
to the shapes of production and death functions and translocation dynamics.
The model is a differential equation model that evolved from experimentalists
who work on marshes interacting directly with project modelers to produce
a working research product. Such model exercises are a continuing endeavor
in the group.
We are developing a model of sediment deposition
on marsh surfaces that will provide quantitative model relationships between
flow events (storms) of various magnitudes and frequencies and resultant
sediment erosion and deposition. We have added a co-PI Wiberg with the
ability to develop this type of model. The model will consist of three
linked models for flow and sediment dynamics in an intertidal marsh environment.
A model will describe sediment deposition from a flow over a vegetated
marsh surface like those at the Mainland Marsh Site. This depositional
model will be linked to a second model that will provide initial flow and
sediment conditions for the marsh deposition model based on flow and sediment
transport conditions in the tidal creeks dissecting the marshes. The third
component model will simulate combined channel and flood plain flow (e.g.
Knight and Demetriou 1983). The conjunction of the three models will provide
an integrated view of flow and sediment dynamics for the VCR LTER marsh
systems. By combining these models with historical data on the frequency
and magnitude of storm events, we will examine the recurrence times and
magnitudes of depositional and erosional hydrodynamic events.
The model
for sediment deposition and erosion on a vegetated marsh is currently being
formulated. Sata and analyses are available from (experimental) studies
of flow resistance due to vegetation (e.g., Turner and Chanmeesri 1984;
Pasche and Rouve 1985; Kadlec 1990). However little has been done to relate
vegetation to erosion, transport, and deposition of sediment. Data to constrain
and test our model of sedimentation on vegetated marsh surfaces will be
collected on a low marsh in the Mainland Marsh Site. Previous work in this
area includes characterization of sediment texture on the marsh surface
and in cores, long-term accumulation rates from lead isotope dating, and
an indication of the level of perennial sediment activity on the low marsh
(Kastler 1993). Future measurements will include flow velocities and suspended
solids associated with tidal flows on natural and lowered vegetated marsh
surfaces (see Table 3).
The VCR LTER has a centralized data management system 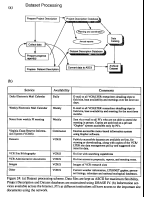 (Fig. 24a) which
is the responsibility of a full-time data manager and a student assistant.
The data manager is a project PI and heads a data management committee.
Our data management philosophy has been that data management begins with
project inception, not project completion, that principle investigators
have primary responsibility for quality assurance, that data should be
accessible by a wide variety of software packages, and that data management
activities should not infringe on the rights of investigators to exploit
the data they collect. A site data management policy has been formulated
and implemented which is based on this philosophy while providing a general
framework for quality assurance and data accessibility.
(Fig. 24a) which
is the responsibility of a full-time data manager and a student assistant.
The data manager is a project PI and heads a data management committee.
Our data management philosophy has been that data management begins with
project inception, not project completion, that principle investigators
have primary responsibility for quality assurance, that data should be
accessible by a wide variety of software packages, and that data management
activities should not infringe on the rights of investigators to exploit
the data they collect. A site data management policy has been formulated
and implemented which is based on this philosophy while providing a general
framework for quality assurance and data accessibility.
The data management
system has four general objectives. The first is to provide archival storage
of data. To accomplish this objective, on-line copies of data files are
backed-up on a variety of tape and optical media with at least one copy
maintained off-site. Data is stored in a standard form (the American Standard
Code for Information Interchange - ASCII) which is almost universally accessible
by different software packages.
A second objective is to provide access
to data by researchers. We facilitate such access using Gopher and World-wide
Web-based on-line information systems. Public data sets are available for
downloading, along with suggested citations crediting specific researchers
and the VCR LTER. The Virginia Coast Reserve Information System (VCRIS)
also includes information on research calendars, current weather forecasts,
personnel and bibliographic citations.
A third objective is to provide
a data catalog. This includes a centralized repository for documentation
and keyword indices that facilitates automated searches. Development of
the catalog entries begins with the initiation of each research project.
An initial project description and yearly progress reports form the basis
of each entry. For individual data sets (a data set consists of observations
containing a common set of variables), additional documentation describes
the storage of the data and specific methodologies used in collecting it.
Thus a hierarchical structure of documentation is formed. The data catalog
is maintained in digital form using a relational database program (currently
DBASE IV).
The final objective is to facilitate linkages between data
sets. To this end, we maintain a number of site-resource databases. These
include a research location database (so that different data sets can use
a common vocabulary of site names), master species lists (which define
a unique 6 letter identifier for each species) and a database of GIS coverages.
Standards are also imposed on the formats for temporal and spatial information
that facilitate merging of different data sets.
Network and Intersite activities of PIs
are tabulated in Table 7.
The funding history for LTER PIs for VCR related projects is presented
in Table 8. In total, research support not provided by the LTER Core grant
or from associated supplemental funding amounted to $2,811, 399 Some of
these funds (e.g. to Galloway-NSF, Dolan-FEMA, and Hayden-NOAA) supported
data collection on the site an addition to VCR data archives. Other funds
went to improve technology used by VCR scientists at the site and in laboratory
development (e.g. Blum-Va Mines, Hayden & Shugart-NASA, Macko-NSF,
and UVa). The majority of the projects went to support question-driven
research at the site. It is noteworthy that $56,000 of the related research
projects came from proposal-based graduate student research and fellowship
awards. As most of the VCR PIs get little or no money for summer stipend
support, these external research projects are often the source of such
stipends. Where the research missions of the external research and the
LTER are compatible this relationship is highly synergistic. The research
staff for many of the related research projects brings new scientists to
the VCR research site. This expansion of the research community is significant
and has resulted in the establishment of ESRA (Eastern Shore Research Association).
This association is being developed with the assistance of the Virginia
Marine Sciences Consortium. It will hold an annual scientific meeting,
maintain an electronic bulletin board and maintain a GOPHER information
system. This association will permit us to fully identify the wider research
community that is interested in the Virginia Coast Reserve and Virginia's
Eastern Shore.
Information from the VCR/LTER has been distributed to individual scientists,
governmental organizations, non-profit organizations and the general public
(Fig. 24b and  Fig. 25). A primary mode of distribution to the scientific
community is via the Virginia Coast Reserve Information System (VCRIS),
an Internet accessible information system using Gopher software. Since
its initiation in the fall of 1992 it has been accessed in over 11,000
individual sessions from over 1,000 individual computers, including access
from Germany, Great Britain, Japan, Austria, Switzerland and Australia.
Data on shoreline change and water quality have been provided to Accomac
and Northampton Counties. We have provided The Nature Conservancy with
up-to-date aerial photography and collaborated with them on the development
of vegetation and bird-nesting maps.
Fig. 25). A primary mode of distribution to the scientific
community is via the Virginia Coast Reserve Information System (VCRIS),
an Internet accessible information system using Gopher software. Since
its initiation in the fall of 1992 it has been accessed in over 11,000
individual sessions from over 1,000 individual computers, including access
from Germany, Great Britain, Japan, Austria, Switzerland and Australia.
Data on shoreline change and water quality have been provided to Accomac
and Northampton Counties. We have provided The Nature Conservancy with
up-to-date aerial photography and collaborated with them on the development
of vegetation and bird-nesting maps.
Communication of research results
to the public has been through several avenues. Brochures describing LTER
research were developed in conjunction with The Nature Conservancy and
the Virginia Museum of Natural History. Articles outlining LTER research
were published in the LTER Network News. Additionally, VCR researchers
(especially Randy Carlson, our site manager) have been active in presenting
talks to local groups. Members of the general public with Internet access
have also been able to use VCRIS.
The VCR LTER maintains a number of inventories. Inventories
perform two basic functions: to facilitate comparative research and to
aid in coordinating extant research. Our research-site database fulfills
both these functions. Maintained in a geographical information system,
it provides researchers with instant access to information on sites used
by present or past projects. Data on sites can be accessed using either
geographical or tabular (type of site, site name) referents. Additionally,
it facilitates merging of disparate data sets by providing a vocabulary
of common site names. Additional inventories include species lists for
avian, mammalian and plant species indexed by island and year and the site
bibliography, listing research publications of the VCR LTER. Archives of
the VCR are summarized in Table 9.
The signatory PIs of the VCR LTER
are Bruce Hayden, John Porter and Herman Shugart. Hayden is the lead PI
and is responsible for project management and administration (Fig. 25).
Other than academic duties, all of Hayden's funded research is focused
on the VCR and his time is fully invested in LTER duties. John Porter is
full time on the LTER program and is fully versed in project management
and administration and can serve in that capacity if called on. Shugart
has served as lead PI and is able to serve if required. John Porter is
the VCR data manager and is in charge of our GIS laboratory. Shugart is
our leader in modeling and synthesis. The signatory PIs have primary responsibility
for network and international activities.
During the academic year
PIs in Charlottesville meet weekly and in the summer biweekly to deal with
general administrative issues, advice and consent regarding policy and
procedures and response to NSF, Network and Intersite activities. The minutes
of these meetings are distributed through e-mail the following day and
archived in the project GOPHER VCRIS which is open to all VCR Scientists.
In academic year 1993, we began monthly project-administrative meetings
with the graduate students. Quarterly, we hold meetings of all PIs. The
location of this meeting varies according to needs. In addition to the
PIs, officials of The Nature Conservancy are invited to attend these meetings.
The winter quarterly meeting is our All VCR Scientists meeting.
At this meeting oral research presentations (including REU presentations)
are made and short written versions are subsequently collected, reprinted
and bound for general distribution.
VCR scientists are classified as
Signatory PIs, Co-PIs, Subcontracting PIs, and Affiliated PIs. Signatory
PIs as indicated on the cover-sheet of this proposal have management and
administrative responsibilities and oversight and coordination of research.
Significantly, these PIs have the additional responsibility to insure project
communication including, timely reports to the National Science Foundation,
annual scientific meetings, intersite and international activities, weekly
and quarterly administrative meetings, and long-term research planning
sessions (Figure 25). Hayden serves as Corresponding PI and is the main
contact with NSF and with the LTER Network Office (Figure 25). Co-PIs
are scientists with a long-term research commitment to the VCR LTER research
program and are responsible for the conduct of the research outlined in
this proposal. Sub-contracting Co-PIs have part of their research financed
by the Core VCR LTER grant through subcontracts with UVa. Affiliated PIs
are scientists whose research is independently funded and have research
missions consistent with the VCR LTER, support our data and reporting protocols
and contribute to the data archives of the project including long term-data
sets. Logistics support for their research is provided where possible.
The scientists in each category are listed in Table 1 following the last
page of the Results of Prior NSF Support section of the proposal.
In
past years we used a committee of 5 outside scientists (Dr. John C. Kraft
, Dr. Robert C. Harris, Dr. F. J. Vernberg, Dr. R. G. Wiegert, and Dr.
T. Webb III) as an oversight committee.We intend to continue to rely on
this committee eventhough we had to rely on substitute on occassion. For
the renewal period, we propose to invite these 5 outside reviewers to our
annual VCR All Scientist meeting and each year invite two reviewers to
evaluate specific LTER activities, e.g. data management, modeling, marsh
productivity, decomposition. In the year before our mid-term review and
in the year before our next renewal we will bring our 5 external reviewers
in for intensive reviews. We intend to invite experts from other LTERs
and LMERs to help us in our internal reviews. These individuals know the
requirements of the LTER program, have served on site review panels, and
NSF LTER renewal panels. Project leadership will select the reviewers asked
in each year to evaluate activities and move research programs forward.
We will ask of these reviewers to submit a written set of recommendations.
In 1994 we will begin
a new long-term data acquisition program to determine water volumes in
Hog Island Bay. The bay is comprised of extensive areas of shallow water,
for which no existing depth profiles are available. Our boats will be rigged
with digital fathometers and GPS units so that systematic bathymetric surveys
can be conducted. These data sets collected will be used to determine long-term
trends in bathymetry and estimate the horizontal fluxes of nutrients and
sediment fluxes. Surveys of opportunity resulting from general boat traffic
will also be added to bathymetric data sets. With this data, water volumes
as a function of tide stage can be determined. Oertel, Porter, Hayden and
Kochel will conduct these surveys.
The shoreline and the seaward margins of terrestrial
vegetation are currently determined by occasional aerial photography. In
1994, we plan to begin mapping these ecotones using kinematic GPS equipment.
The resolution of these new data (< 1m) will be better than possible
using photographs (2 m) and can be done on short notice. This kinematic
survey capability will permit accurate mapping of ground-visible ecotones
and critical landscape margins. Kinematic surveys will be repeated from
time to time to determine long term trends in ecotone positions. Hayden,
Porter, Dolan and Kochel will conduct this work.
Animal exclosures are planned
for the Summer and Fall of 1994. Exclosures will be placed at 4 locations
along the North Hog Chronosequence. The exclosures will be placed on 10
yr., 35, yr., 70 yr. and 120 yr landscapes. Exclosures will be 5 m on a
side and will be designed to exclude deer, rabbits and muskrats. Young,
Dueser, Porter, and Moncrief will conduct these experiments.
In 1994, a nitrogen budget
project will be started on Hog Island. The sites for this study are a series
of vegetated pimples some of which have nitrogen fixing Myrica vegetation
and others have no nitrogen fixers. An NADP station is located within a
mile of these pimples and nitrogen fixing and dry and wet deposition data
will be available. There are no runoff losses or losses to groundwater
that are not recoverable. Galloway, Young, and Macko will conduct this
project.
Supplemental
support for the period 1988-1994 was used to 1) support undergraduate students
(REU program), 2) develop technical capabilities and meet LTER network
standards for emerging technologies (Technical Supplements), and 3) meet
special research opportunities (SGER). The specific history of our supplemental
support is given in Table 10.
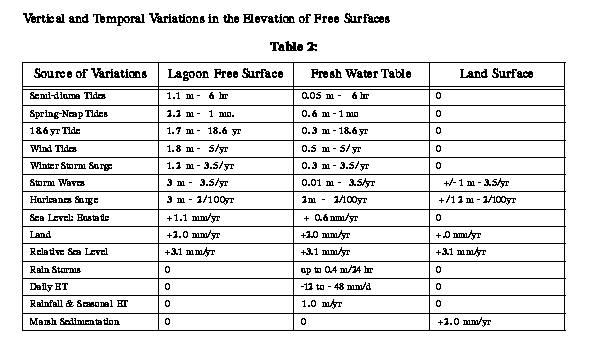{kind=link}
{kind=link}
{kind=link}
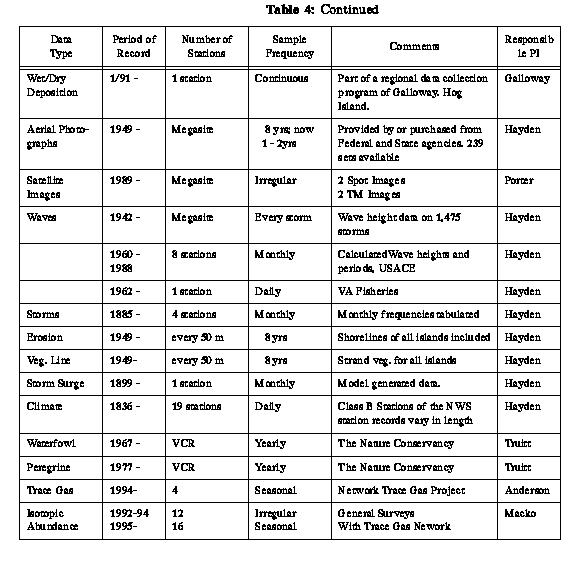{kind=link}
{kind=link}
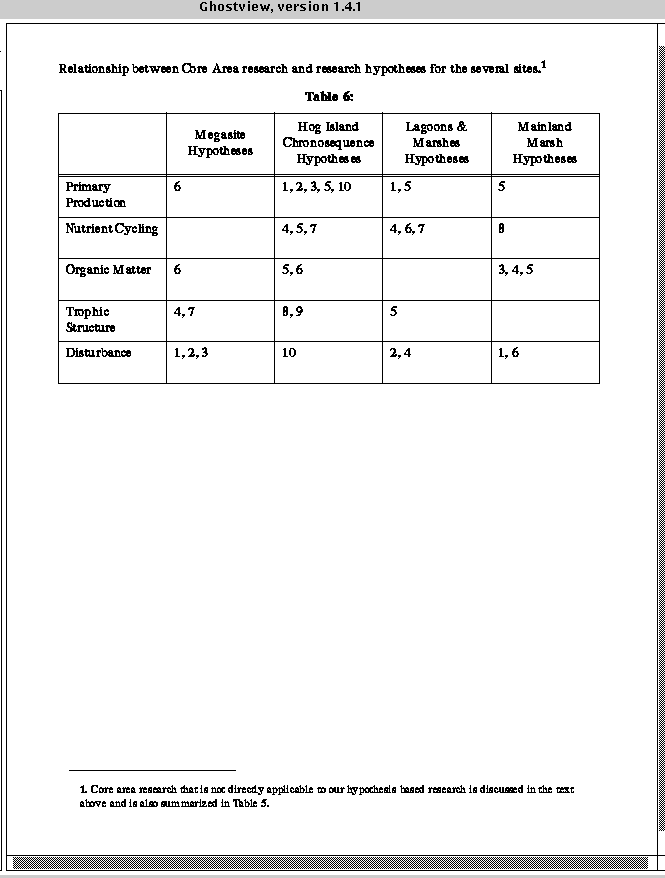{kind=link}
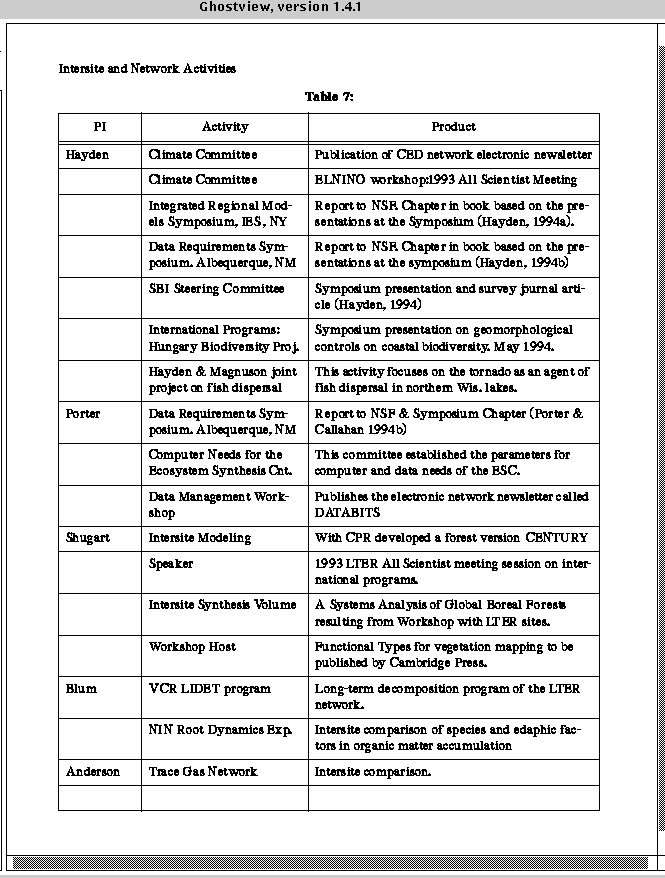{kind=link}
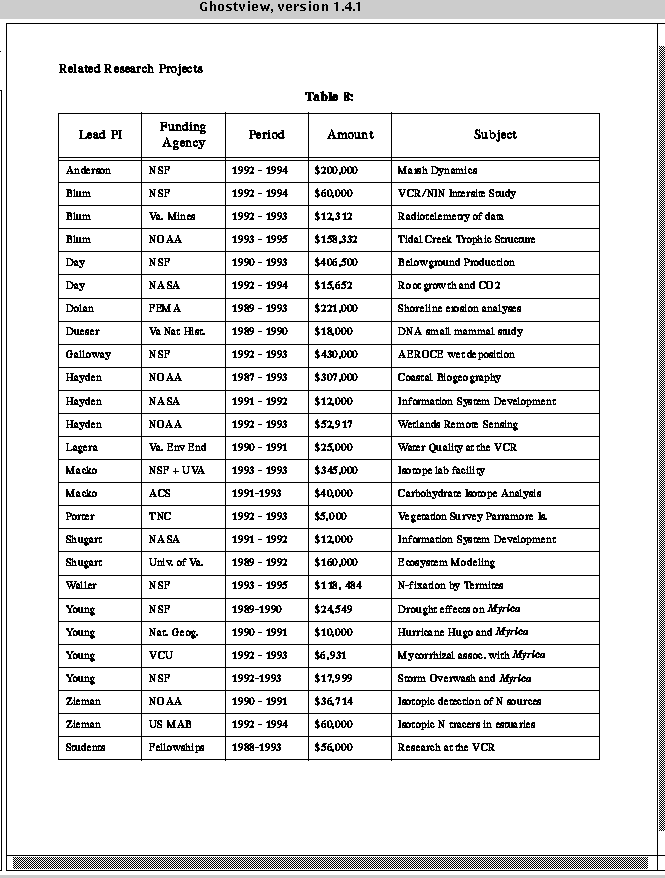{kind=link}
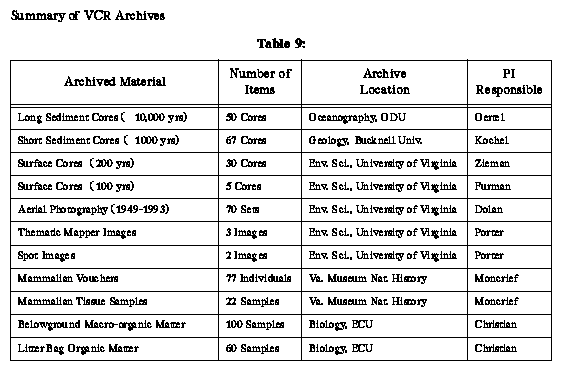{kind=link}
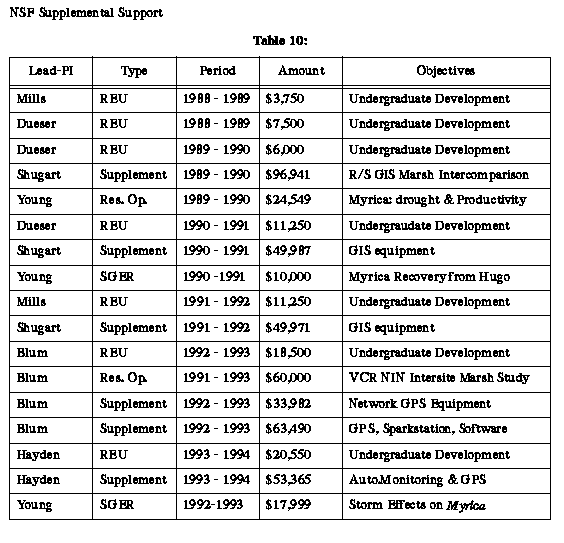{kind=link}